 2019, Vol. 35
2019, Vol. 35中国科学院微生物研究所、中国微生物学会主办
文章信息
- 曲媛媛, 戴春晓, 张旭旺, 马桥
- Qu Yuanyuan, Dai Chunxiao, Zhang Xuwang, Ma Qiao
- 吲哚——种间及跨界信号分子新成员
- A new interspecies and interkingdom signaling molecule—Indole
- 生物工程学报, 2019, 35(11): 2177-2188
- Chinese Journal of Biotechnology, 2019, 35(11): 2177-2188
- 10.13345/j.cjb.190158
-
文章历史
- Received: April 25, 2019
- Accepted: September 2, 2019

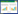
引用本文

|



2. 大连理工大学 食品与环境学院 工业生态与环境工程教育部重点实验室,辽宁 盘锦 124221;
3. 大连海事大学 环境科学与工程学院 环境系统生物学研究所,辽宁 大连 116026
2. Key Laboratory of Industrial Ecology and Environmental Engineering (Ministry of Education), School of Food and Environment, Dalian University of Technology, Panjin 124221, Liaoning, China;
3. Institute of Environmental Systems Biology, College of Environmental Science and Engineering, Dalian Maritime University, Dalian 116026, Liaoning, China
作为一种典型的含氮杂环芳烃化合物,吲哚在自然界中广泛存在。微生物与动植物的生理生化过程会伴随着吲哚的产生,研究表明,包括大肠杆菌Escherichia coli、霍乱弧菌Vibrio cholerae在内的约有150种细菌能够在体内色氨酸酶的作用下合成吲哚,动物肠道中吲哚浓度可高达1.1 mmol/L[1]。然而越来越多的研究表明,吲哚不仅仅是单纯的氮杂环化合物或代谢物,更是一种新型种间及跨界的信号分子。自然生境中吲哚的广泛存在使得其生物学意义及归趋受到广泛关注,成为环境保护、生态平衡和人类健康等诸多领域的热点化合物。
近年来,研究人员证明了吲哚在微生物代谢、人类健康及植物生长等多个方面扮演着重要角色。吲哚能够调节微生物的生长状态与存活率,影响细菌的耐药性、病原菌毒性、孢子形成和细胞分裂等多种生理生化行为,并对微生物群落中生物膜的形成、群感效应以及极端环境适应能力等生物功能有显著影响[2]。对于植物而言,吲哚与其生长发育和防御系统的形成过程密切相关[3]。吲哚也与人类常见疾病如肠道炎症、神经系统疾病及糖尿病等密切相关,并能够调控动物细胞的氧化压力以及荷尔蒙的分泌[4-5]。不难看出,吲哚在自然生境中具有生物学与生态学双重意义,其生物代谢及信号机制成为环境微生物学、生态学以及人体健康等交叉领域的研究热点与难点。本文将重点对吲哚作为新型种内、种间及跨界信号分子的相关研究进展进行综述,并对该领域的研究方向进行展望。
1 从生物代谢到信号分子的研究历史吲哚在自然界中广泛存在,最早可以追溯到1897年,美国病理学家Theobald Smith发现大肠杆菌E. coli和霍乱弧菌V. cholerae可以合成大量吲哚[6],这一现象引起了学者的持续关注。Wood等在1947年提出吲哚是微生物代谢色氨酸的代谢产物[7]。而后1972年,Watanabe等提出这一过程经由微生物体内的色氨酸酶TnaA所调控[8],即色氨酸在微生物体内由TnaA代谢为吲哚、丙酮酸和氨。代谢得到的吲哚分子由于其具有疏水性,可自由扩散通过细胞膜,或由Mtr或AcrEF蛋白转运至微生物体外[9]。截止2015年,共有150种革兰氏阳性菌和革氏兰阴性菌被报道可以产生吲哚[1],但目前尚未发现真菌可以合成吲哚。
作为一种典型的含氮杂环芳烃化合物,吲哚被广泛应用于医药、染料及各类化工行业中,在焦化废水以及畜牧废水等难降解废水中大量存在,并会通过各种工业废水或废物的排放进入到自然环境中[10]。因此在环境领域,吲哚的研究主要集中于其作为含氮杂环化合物的微生物降解途径。自1928年Gray首次报道氧化吲哚假单孢菌Pseudomonas indoloxidans降解吲哚以来[11],研究者们对不同菌株降解吲哚的化学途径进行了探索,虽然降解途径各有差异,但大都遵循吡咯环氧化开环途径。迄今,已报道的吲哚降解途径主要有3种,即邻苯二酚途径、邻氨基苯甲酸途径及龙胆酸途径[12]。笔者团队关于吲哚降解及资源化开展了数年的研究,分别筛选出了10余株具有吲哚降解能力的菌株,对苯酚羟化酶、萘双加氧酶、吲哚加氧酶等进行了深入研究,并构建了吲哚资源化体系[12-15]。2017年,笔者团队首次证实菌株贪铜杆菌Cupriavidus sp. SHE降解吲哚遵循好氧代谢中比较罕见的辅酶A途径,而非双加氧酶和龙胆酸代谢途径[13]。吲哚转化方面,研究表明一些常见的芳烃代谢加氧酶如萘双加氧酶、细胞色素P450加氧酶及黄素单加氧酶等,会氧化吲哚生成靛蓝、异靛蓝、靛玉红等高附加值产物[14-16]。然而,长期以来研究者们一直关注着吲哚在自然环境中的迁移转化,而忽视了代谢过程的主宰者——原核或真核生物,是否也同时受到吲哚的反作用。研究表明,芳烃代谢途径是代谢交换因子的重要资源,在微生物与微生物、微生物与动植物相互作用关系中发挥着前所未知的生物学作用[17]。不难看出吲哚的种间或跨界信号作用机制将成为新的研究增长点,具有重要的理论及实际意义。因此,揭示基于吲哚的“合成 & 降解 & 转化”的物质交换过程的生态机制,对于生态学及环境污染控制领域都是极其关键的。
虽然微生物合成或降解的现象已经被发现超过了100多年,但是吲哚这一分子的实际生物学作用在近20年才逐渐被深入研究。1993年Gerth等首次提出吲哚是橙色标桩菌Stigmatella aurantiaca的自诱导剂后[18],许多研究小组逐渐开始对吲哚的多种生物学功能进行研究。1999年,吲哚首次被认为是大肠杆菌潜在的自诱导剂[19],紧接着在2001年,吲哚被确认是一种能够激活astD、gabT和tnaB等基因的细胞间信号分子[20]。研究初期,关于吲哚是否为细胞间信号分子这一观点,在很长一段时间内存在争议,然而,更多的研究证明了吲哚满足了细胞-细胞信号分子的4个标准[21-22]:1)吲哚主要在对数生长期产生,在稳定期大量累积[20];2)大肠杆菌等微生物能在体外累积,且尽管吲哚具有疏水性,能扩散穿过细胞膜̼但研究证明AcrEF、Mtr蛋白作为特定的识别器分别负责了吲哚的进入与排出[9];3)吲哚可调控细胞分裂等生理活动,且不同浓度吲哚影响效果不同[23];4)吲哚产生的信号引起了超出代谢或解毒信号所需的生理变化,也会对吲哚降解菌生物膜的形成与细胞分裂等生理活动产生影响[24]。吲哚作为新型种内、种间及跨界信号分子,在微生物及其群落间相互作用机制方面的研究具有重要意义。
2 吲哚是种内及种间信号分子 2.1 吲哚调节微生物的生理状态吲哚作为信号分子,能够控制微生物的多种生理功能,如提升质粒稳定性[25]、参与细胞的生长与分裂过程[26-27]、影响抗生素耐受性[28-30]及细胞膜的形成[26, 31-33]等,而这些研究主要集中在产吲哚菌株E. coli和V. cholerae中(表 1)。此外近年来,吲哚也被证明可以增强E. coli在恶劣条件中的存活率。Lai等证明了E. coli在厌氧条件下能产生吲哚,降低了硝酸盐厌氧呼吸过程中代谢物的毒性[34]。Liu等则发现外加低浓度的吲哚能够提升敲除色氨酸酶的E. coli在高温条件下的存活率[35]。除了E. coli和V. cholerae以外,吲哚也可以影响其他产吲哚菌,如抑制蜂房芽胞杆菌Paenibacillus alvei和橙色标桩菌Stigmatella aurantiaca的耐药性孢子的形成[36],提升植物内生病原菌Pantoea ananatis在营养匮乏条件中的存活率[37]等。在自然环境下的复杂微生物群落中,微生物之间能够相互妨碍或利用对方产生的信号分子。研究证明,E. coli分泌的大量吲哚会被其他菌株如洋葱伯克霍尔德氏菌Burkholderia cepacian G4羟化生成羟基吲哚,且反向调控E. coli生物膜的形成[26]。由此可见,吲哚对产生吲哚产生菌和非吲哚产生菌的生理作用可能是完全不同的。
| 现象 | 作用机制 | 文献 | |
| 生物膜 | 吲哚增强菌株生物膜形成 | 色氨酸酶缺失降低生物膜形成能力 | [24] |
| 吲哚抑制生物膜的形成 | 影响调控群感响应蛋白SdiA进而影响生物膜 | [26] | |
| 通过多元胁迫抗性蛋白YcfR诱导吲哚合成及生物膜的形成 | [32] | ||
| 热激蛋白IbpAB导致的内源性氧化胁迫会提高吲哚生成 | [33] | ||
| 生物膜调控基因bssR和bssS参与了吲哚的跨膜运输 | [31] | ||
| 生长 | 高浓度吲哚可逆的抑制了细胞的分裂 | 调控SdiA,抑制细胞分裂启动子ftsQ2p的表达 | [26-27] |
| 质粒稳定性 | 吲哚能够通过调节细胞分裂调节子提高质粒的稳定性 | [25] | |
| 存活率 | 耐酸性 | 提高耐酸系统(谷氨酸脱羧酶)相关基因的表达 | [26] |
| 厌氧硝酸盐呼吸 | 吲哚和呼吸产生代谢物反应生成硝基吲哚衍生物 | [34] | |
| 高温条件 | 温度影响了tnaA基因表达,提升了在50 oC条件下的存活率 | [35] | |
| 耐药性 | 上调多药外排泵的相关基因的表达,提高菌株对多种抗生素的耐药性 | 通过双信号调控系统BaesR/CpxAR | [28] |
| 激活转录因子GadX | [29] | ||
| 通过SdiA调控多药流出泵基因 | [30] |
非吲哚产生菌也能够响应产吲哚菌分泌的吲哚分子,一些非吲哚产生菌则会编码一些加氧酶进一步降解或者转化吲哚生成吲哚同系物。此外,吲哚也控制了非吲哚产生菌的多种生理活动(表 2),如抑制细菌群感效应(QS),影响毒性蛋白的表达及多种菌株之间的粘合因子等[38-46]。吲哚能够影响群落生物膜的生成,如能够抑制由固着生物膜形成的蓝藻水华[43],减少绿针假单胞菌Pseudomonas chlororaphis的生物膜形成[37]等。除此之外,吲哚也会调整其他不产吲哚菌的抗生素耐受性,如铜绿假单胞菌Pseudomonas aeruginosa、恶臭假单胞菌Pseudomonas putida、根癌农杆菌Agrobacterium tumefaciens等[23, 38, 45-46]。吲哚也可以通过抑制细胞黏着行为和抑制毒素产生来降低不产吲哚病原菌的毒性,例如,吲哚被证明能够阻止真菌病原体白色念珠菌Candida albicans黏附在线虫肠道上皮细胞[42],吲哚还能够减少线虫体内的金黄色葡萄球菌Staphylococcus aureus表达毒性色素[44]。尽管如此,关于吲哚对于非产吲哚菌的生理影响的研究尚不够全面,有待进一步的探讨。
| 主要作用 | 作用机制 | 菌属 |
| 群感效应(QS) | 抑制了群感信号分子AHL-调节的色素沉着,从而抑制了群感效应 | Chromobacterium violaceum[38] |
| Pseudomonas chlororaphis[38] | ||
| Serratia marcescens[38] | ||
| 抑制了群感效应,进而抑制了生物膜的生成 | Pseudomonas aeruginosa[39] | |
| 通过抑制QS调节蛋白的折叠,抑制了生物膜的生成 | Acinetobacter oleivorans[40] | |
| 吲哚会上调群感淬灭酶Moml的表达 | Muricauda olearia[41] | |
| 生物膜 | 调节膜、流出泵等相关基因的表达,促进生物膜的生成 | Agrobacterium tumefaciens[23] |
| 通过刺激转录因子,诱导生物膜的形成 | Candida albicans[42] | |
| 形成了固着生物膜,抑制了蓝藻暴发 | Microsystis aeruginosa[43] | |
| 诱导细胞Ca2+外排泵,抑制生长以及生物膜的形成 | Pseudomonas chlororaphis[38] | |
| 毒性因子 | 通过刺激转录因子,诱导毒性的产生 | Candida albicans[42] |
| 葡萄球菌黄素和溶血素的减少,导致了毒性的减少 | Staphylococcus aureus[44] | |
| 抗生素耐受性 | 对基因进行调节,促进抗生素耐受性 | Agrobacterium tumefaciens[23] |
| 调节流出泵的活性,提升抗生素的耐受性 | Pseudomonas putida[45] | |
| Pseudomonas aeruginosa[39] | ||
| 氧化应激反应及外排泵活性提升了抗生素的耐受性 | Salmonella typhimurium[46] |
然而大量的实验证明,吲哚的生理学作用与不同的实验对象以及环境条件息息相关:E. coli S17-1 tnaA突变体减少了生物膜的形成,而添加吲哚后菌株则恢复了菌株生物膜形成[47]。相反的是,吲哚会减少9种非致病性E. coli (BW25113、BW25113 bhsA、BW25113 bssR、BW25113 bssS、BW25113 tnaA、ATCC25404、JM109、TG1和XL1-Blue)以及致病性E. coli O157: H7生物膜形成[26]。在不同实验条件下,吲哚生理功能的矛盾性有可能是归因于研究中使用的各种实验条件的差异性。Mond等提出信号网络应当适应群落的发展程度,即信号分子吲哚的产生应当对整个微生物群落有益[48]。因此,可以推测产生吲哚的细菌可能会使用吲哚来抵抗其他原核或真核生物,而非吲哚产生菌则通过降解吲哚抵御这种影响。吲哚作为细胞-细胞间重要的信号分子,对于吲哚产生菌和非吲哚产生菌的作用机理和作用结果可能都是有区别的。因此,吲哚在复杂微生物群落的活动中扮演的角色值得进一步探索。
2.2 吲哚的生理学作用机制在E. coli中的转运蛋白Mtr以及外排蛋白AcrEF-TolC参与了吲哚的跨膜运输[21]。而吲哚是一种疏水性物质,吲哚穿越细胞的行为也可以不在转运蛋白的参与下自发完成。当吲哚跨越细胞膜时,会导致细胞膜电位的变化,从而进一步发挥调节作用。信号分子的调节方式有两种,分别为持久性和脉冲信号。E. coli在细胞分裂或生长时,其体内会产生吲哚浓度的短暂升高,即瞬时脉冲[26, 49]。这一现象说明,吲哚是一种离子载体,高浓度的吲哚会通过调节细胞膜的电势阻止E. coli细胞的分裂,从而进行应激性的保护。此外,吲哚对于细胞膜、质粒稳定性和酸性条件耐受性等多方面的影响使得吲哚能够调节病原菌的感染过程。吲哚被证明了能够调节病原菌的毒性蛋白的表达,如吲哚通过下调P. aeruginosa的毒性相关基因的表达[39];与之相反,吲哚会促进出血性大肠杆菌毒性相关蛋白(如EspAB)的分泌[50]。而对于更多的致病菌,吲哚的调控作用与机制还需要进一步的研究与完善。
吲哚分别从药物外排泵及持留菌的生成两方面对菌株的抗生素耐受性进行调节。首先,吲哚能够通过影响大肠杆菌的药品流出泵及抗氧化保护机制从而对大肠杆菌的抗生素耐受性进行调整。吲哚可以通过影响两种物质信号传导系统(BaeSR和CpxAR),进而诱导大肠杆菌异型生物质外排基因的表达[27, 50],大肠杆菌也会在抗生素存在的条件下产生更多的吲哚。吲哚诱导了鼠伤寒沙门氏菌Salmonella typhimurium的外排泵系统(AcrAB)的转录调节器RamA,进而提高了菌株耐药性[46]。此外,吲哚被证明能够增多大肠杆菌耐受抗生素的持留菌的形成[51]。然而,相反的是,它也被报道过能够通过影响磷酸二酯酶DosP降低了环磷酸腺苷浓度,以及增多表达降解色氨酸酶所表达的mRNA的毒性蛋白YafQ,减少了TnaA蛋白的表达,进而减少耐药性持留菌的形成[52]。
2.3 吲哚是一种新型群感效应信号分子吲哚作为信号分子的作用更加明显体现在吲哚对于群体感应(QS)的调控作用。群感效应指微生物能够感知种群大小并相应调节对应生理活动的能力,而细菌则会生产多种群感效应分子,如酰基高丝氨酸内酯(AHLs)、自体诱导物Ⅱ类分子(AI-2)等,并利用这些分子来影响孢子形成、生物体发光、毒性因子的产生等。与AHL的群感效应调节功能正相反,吲哚能够作为AHL信号分子群感效应淬灭剂,在微生物中广泛传播。研究证明,0.5–1.0 mmol/L吲哚便可破坏紫色色杆菌Chromobacterium violaceum、粘质沙雷氏菌Serratia marcescens、铜绿假单胞菌P. aeruginosa和绿针假单胞菌P. chlororaphis中的群感效应[38]。吲哚会使菌株中群感效应控制基因的差异表达,如吲哚引起了许多分子伴侣和蛋白酶编码基因上调使得AqsR蛋白折叠受到抑制[40]。除此以外,吲哚也被证明会与群感效应响应蛋白相互影响。SdiA是大肠杆菌中可与群感信号分子AHL相互作用的响应蛋白。研究表明,在大肠杆菌中基于吲哚和基于AHL的信号传导之间会相互影响。虽然SdiA活性已经被证明会被E. coli产生的吲哚所抑制[26],但没有直接证据表明吲哚可以与任何SdiA同源物(或SdiA本身)结合。另外,能够与AHL结合的TraR蛋白可以不受吲哚影响,但不含AHL的TraR蛋白在吲哚存在下发生错误折叠[53]。这表明群感效应调节器需要同源信号来稳定,但它们的折叠会受到吲哚的影响,从而进一步影响群感效应。尽管如此,吲哚的受体系统以及该化合物作为信号分子在微生物及群体协调行为中的作用尚需要进一步证实。
研究证明,吲哚和AI-2信号分子的作用机制相仿,但作用往往截然相反[18]。吲哚和AI-2均是较为少见的能够对革兰氏阳性和革兰氏阴性细菌共用信号分子。与AHL系统不同,目前尚未发现直接与吲哚或AI-2结合的LuxR型蛋白,受体系统需要进一步研究。大肠杆菌在趋化实验中趋向于AI-2信号分子,而受到吲哚的排斥[54-55]。AI-2能够增加大肠杆菌的运动性,而吲哚则会减弱大肠杆菌的运动性[54-55]。在37 ℃,AI-2发挥的基因调控作用更加显著,而吲哚最优作用温度则是30 ℃[56]。探索AI-2和吲哚两种信号分子如何在自然环境中的作用机制是极其重要的。
在环境污染控制领域,吲哚长期被认为是一种典型的氮杂环污染物,研究大多集中于污染物降解方面,而在环境生物技术中的实际应用尚且欠缺。近年来,群感效应在环境水处理生物反应器中的作用受到越来越多的关注[57]。而吲哚能够干扰典型群感效应信号分子AHL的表达及作用,又与AI-2分子有着相似的基础性质和不同的作用效果,被认为是一种潜在的群感效应淬灭剂(QSI)[2]。在水处理环境工程中,吲哚是否能与AHL分子以及AI-2分子进行联合调节环境工程中的微生物群落,从而应用于膜污染控制、污泥性能调节等水处理工程中,是值得验证的。
3 吲哚的跨界信号调控 3.1 吲哚跨界信号调控植物与微生物、动物间的相互作用吲哚是典型植物次级代谢产物之一,天然花油中就含有一定量的吲哚。植物体内合成吲哚最为常见的途径是莽草酸途径[12]。莽草酸在植物体内的莽草酸激酶(KS)作用下,生成3-吲哚磷酸莽草酸,经过系列酶促反应后被代谢成为3-吲哚磷酸甘油,最终3-吲哚磷酸甘油被色氨酸合成酶(TSA)催化为吲哚。吲哚进一步被植物体内的色氨酸合成酶(TSB)生成L-色氨酸,在植物体内被催化为各种吲哚生物碱,调控植物的各种生理活动。此外,部分植物会释放多种吲哚等挥发性有机物来吸引植食性昆虫的天敌或寄生蜂,以免受侵袭(图 1)。草食性动物的攻击会刺激玉米中磷酸盐裂解酶的表达,促进植物体内吲哚-3-甘油及吲哚的排放,进而启动一系列的防御体系[58]。植物释放的吲哚不仅仅会对自身起调节作用,更会作用于周围的同种或异种植物,引起植物挥发性萜烯的释放和茉莉酸(JA)、脱落酸(ABA)的产生,增强植物对外来害虫与真菌的抵抗力[59]。

|
| 图 1 吲哚作为跨界信号分子在生态环境中起到的作用 Fig. 1 Functions of indole as interkingdom signal molecule in nature.①: indoles produced by intestinal microorganism influence the physiological behavior of animals or human health; ②: indole producing bacteria and indole non-producing bacteria exchange metabolites in microbial communities; ③: there are indoles in plants and rhizobacteria which influence the growth and defence behavior of plants; ④: indoles produced by plants will modulate the behavior of insects and animals. |
| |
土壤微生物产生及降解吲哚的一系列行为也对植物的生长及根系发育有着一系列的调控作用(图 1)。根际微生物产生的吲哚也能够剂量依赖性地提升白菜幼苗的活力指数和鲜重[60]。吲哚通过与植物生长素、细胞分裂素和油菜素类固醇激素通路相互作用,从而刺激拟南芥的生长[3]。除了吲哚以外,微生物会合成一系列吲哚衍生物,如吲哚-3-乙酸、吲哚-3-丁酸等,均为常见的植物生长调节剂[61]。其中,吲哚-3-乙酸不仅仅会影响寄主植物生理活动,也可以整合进入细菌复杂代谢网络,调节细菌自身基因表达与生理代谢,作为跨界信号参与植物与微生物的相互作用[62]。不同吲哚衍生物可能对植物生长有着不同的影响,因此进一步了解吲哚及其衍生物在根际微生物和植物中的调节作用,有助于进一步揭示植物-微生物相互作用关系。
3.2 肠道微生物产生的吲哚参与了动物生理调节动物的汗液、尿液以及排泄物中均含有较高浓度吲哚,如人体消化道和排泄物中的吲哚浓度可达0.25–1.1 mmol/L,是动物肠道和排泄物中臭味的主要来源之一,这是因为动物体肠道中含有大量产吲哚的肠道微生物(图 1)。代谢组学研究表明,肠道内微生物的存在决定了哺乳动物循环系统中吲哚类物质的存在,并参与到肠道共生菌群的建立过程中,进而影响如病菌感染、肠道炎症和糖尿病等人类疾病[4]。此外,吲哚能使肠道上皮细胞联结更加紧密,并增加细胞抗炎因子在人体HCT-8细胞中的分布[63]。肠道微生物产生的吲哚甚至会对小鼠的大脑兴奋性产生负面影响[64]。肠道微生物也能够通过产生吲哚,影响宿主的内分泌L细胞,调节肠促胰岛素肽GLP-1,进而影响宿主的生理代谢活动[65]。同时,吲哚也被认为是一种芳烃受体(AhR)的自然配体和激活物质[66],AhR是人类体内由配体激活的转录因子,参与调节人体中芳香烃物质的生物反应,从而影响体内免疫响应并调节多种异生素代谢酶(如细胞色素P450等)的表达。肠道微生物还会将吲哚转化成羟基吲哚、甲基吲哚、硫酸吲哚酚和靛红等各种吲哚衍生物,这些物质在动物血液、周围组织、尿液甚至脑组织中的浓度可达到0.01–0.2 mmol/L[2]。2018年,Mathilde等的研究证明,肠道微生物产生的吲哚进而被氧化生成神经抑制剂靛红、羟化吲哚等衍生物积累于小鼠大脑,导致了运动活性的急剧下降。研究者同时比对了人类微生物宏基因组数据库中的色氨酸酶,得出体内肠道微生物倾向于产吲哚的人更易于产生焦虑和情绪障碍的结论[64]。人体肝性脑病和帕金森疾病往往伴随着血液中的氧化吲哚及尿液中靛红的含量的下降[67-68]。吲哚-3-乙酸则被认为能够减弱肝细胞中细胞激素调节的脂肪的形成,对非酒精性脂肪肝有着潜在的有益调节作用[69]。肠道微生物代谢产物吲哚丙酸则被认为是Ⅱ型糖尿病发展的潜在生物标记物[70]。可见,吲哚类物质与人体健康有着十分密切的联系,仍需要深入研究。
吲哚及其衍生物也会直接影响昆虫的行为,如5-碘代吲哚会导致线虫体内细胞巨大化而死亡[71]。除此之外,昆虫如蚊子和蝴蝶等,也能够识别微生物或植物产生的吲哚,由于吲哚的独特气味,吲哚的受体在这些物种之间广泛存在(图 1)。吲哚来源于色氨酸的代谢过程,因此吲哚对于各界的物种来说是一种营养物质色氨酸存在位置的提示,动物可以基于吲哚进行代谢调整[2]。昆虫会受到吲哚气味的吸引,线虫会更趋向于捕食产吲哚菌[72]。可见,微生物代谢吲哚并转化生成吲哚衍生物对于动物的生理代谢功能有着极为重要的作用,仍需进一步的探究。
4 展望随着吲哚的研究逐渐从作为典型污染物的生物降解向微生物种间及微生物-动植物间跨界信号传导机制研究转变,吲哚的生理学及生态学的意义受到越来越多的关注。吲哚不仅仅单纯参与了细菌的耐药性、病原菌毒性、细胞分裂及极端环境适应能力等多种生理生化行为,更是微生物群落中整体调控的一种潜在信号分子[73]。吲哚也能够调整动物的生理行为与健康状态,影响植物的生长与防御机制,在自然系统中有着无可替代的价值[62, 64]。对于吲哚跨界信号调控机制的研究意味着信号传导打破了种属的限制,更多关注于自然系统的整体信号调节作用。
今后的研究中可以关注以下几个问题:1)关于吲哚的现有研究大多集中于产吲哚菌、两株菌之间、或单一微生物与昆虫之间的简单体系,而将这类新型信号分子置于复杂环境中的研究较少;由于吲哚在自然系统中的广泛存在,吲哚能够对自然系统中的每一个子系统发挥信号作用,然而仍缺少对于吲哚跨界发挥作用的完整论述,将吲哚与其他信号分子相联系起来,探究多种信号分子在体系中的协同与拮抗作用,有助于解析信号分子在生态环境中发挥复杂作用。2)吲哚本身就是多种药物的核心骨架,且吲哚能够调节共生菌群的各方面生理状态,进而能够对动物健康产生影响,可见吲哚在肠道共生菌群的研究中不可忽略,但现有的研究尚不足。将吲哚类跨界信号分子对共生菌群建立的影响与人体健康相结合,可以更好地揭示肠道共生菌群与人体健康之间的有机联系。3)此外,作为新型信号分子,吲哚在实际中的应用有待开发。吲哚具有群感效应淬灭的作用,能够减少生物膜的形成,有代替抗生素作为抗菌剂的潜力。此外,吲哚作为一种群感效应相关的信号分子,在水处理领域对于减少膜污染、调节颗粒污泥的形成过程、调节活性污泥的性能等方面值得进一步的探讨。
可见,从更多方面对吲哚信号分子的生理学作用进行考察,同时对复杂环境中吲哚信号分子的产生-传递-降解过程深入讨论,将有助于解析吲哚类物质的生态学意义及其在生物工程领域的潜在应用价值。
| [1] |
Han Y, Sun MM, Wang JP, et al. Progress on indole: an intercellular signal molecule in microbial communities. Microbiol China, 2015, 42(4): 736-748 (in Chinese). 韩茵, 孙苗苗, 王建平, 等. 吲哚作为细菌细胞间信号分子的研究进展. 微生物学通报, 2015, 42(4): 736-748. |
| [2] |
Lee JH, Wood TK, Lee J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol, 2015, 23(11): 707-718. DOI:10.1016/j.tim.2015.08.001 |
| [3] |
Sharifi R, Lee SM, Ryu CM. Microbe-induced plant volatiles. New Phytol, 2018, 220(3): 684-691. DOI:10.1111/nph.14955 |
| [4] |
Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci, 2017, 20(2): 145-155. DOI:10.1038/nn.4476 |
| [5] |
Kennedy PJ, Cryan JF, Dinan TG, et al. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology, 2017, 112: 399-412. DOI:10.1016/j.neuropharm.2016.07.002 |
| [6] |
Smith T. A modification of the method for determining the production of indol by bacteria. J Exp Med, 1897, 2(5): 543-547. DOI:10.1084/jem.2.5.543 |
| [7] |
Wood WA, Gunsalus IC, Umbreit WW. Function of pyridoxal phosphate: resolution and purification of the tryptophanase enzyme of Escherichia coli. J Biol Chem, 1947, 170: 313-321. |
| [8] |
Watanabe T, Snell EE. Reversibility of the tryptophanase reaction: synthesis of tryptophan from indole, pyruvate, and ammonia. Proc Natl Acad Sci USA, 1972, 69(5): 1086-1090. DOI:10.1073/pnas.69.5.1086 |
| [9] |
Yanofsky C, Horn V, Gollnick P. Physiological studies of tryptophan transport and tryptophanase operon induction in Escherichia coli. J Bacteriol, 1991, 173(19): 6009-6017. DOI:10.1128/jb.173.19.6009-6017.1991 |
| [10] |
Huang Y, Hou XL, Liu ST, et al. Correspondence analysis of bio-refractory compounds degradation and microbiological community distribution in anaerobic filter for coking wastewater treatment. Chem Eng J, 2016, 304: 864-872. DOI:10.1016/j.cej.2016.05.142 |
| [11] |
Gray PHH. The formation of indigotin from indol by soil bacteria. Proc Roy Soc B: Biol Sci, 1928, 102(717): 263-280. DOI:10.1098/rspb.1928.0003 |
| [12] |
Ma Q, Zhang XW, Qu YY. Biodegradation and biotransformation of indole: advances and perspectives. Front Microbiol, 2018, 9: 2625. DOI:10.3389/fmicb.2018.02625 |
| [13] |
Qu YY, Ma Q, Liu ZY, et al. Unveiling the biotransformation mechanism of indole in a Cupriavidus sp. strain. Mol Microbiol, 2017, 106(6): 905-918. DOI:10.1111/mmi.13852 |
| [14] |
Zhang XW, Qu YY, Ma Q, et al. Illumina MiSeq sequencing reveals diverse microbial communities of activated sludge systems stimulated by different aromatics for indigo biosynthesis from indole. PLoS ONE, 2015, 10(4): e0125732. DOI:10.1371/journal.pone.0125732 |
| [15] |
Zhang XW, Qu YY, Ma Q, et al. Cloning and expression of naphthalene dioxygenase genes from Comamonas sp. MQ for indigoids production. Process Biochem, 2013, 48(4): 581-587. DOI:10.1016/j.procbio.2013.02.008 |
| [16] |
Ma Q, Qu YY, Zhang XW, et al. Recent advances in microbial synthesis of indigo. Chin J Appl Environ Biol, 2012, 18(2): 344-350 (in Chinese). 马桥, 曲媛媛, 张旭旺, 等. 靛蓝的微生物合成研究新进展. 应用与环境生物学报, 2012, 18(2): 344-350. |
| [17] |
Díaz E, Jiménez JI, Nogales J. Aerobic degradation of aromatic compounds. Curr Opin Biotech, 2013, 24(3): 431-442. DOI:10.1016/j.copbio.2012.10.010 |
| [18] |
Gerth K, Metzger R, Reichenbach H. Induction of myxospores in Stigmatella aurantiaca (myxobacteria): inducers and inhibitors of myxospore formation, and mutants with a changed sporulation behaviour. J Gen Microbiol, 1993, 139(4): 865-871. DOI:10.1099/00221287-139-4-865 |
| [19] |
Baca-DeLancey RR, South MMT, Ding XD, et al. Escherichia coli genes regulated by cell-to-cell signaling. Proc Natl Acad Sci USA, 1999, 96(8): 4610-4614. DOI:10.1073/pnas.96.8.4610 |
| [20] |
Wang DD, Ding XD, Rather PN. Indole can act as an extracellular signal in Escherichia coli. J Bacteriol, 2001, 183(14): 4210-4216. DOI:10.1128/JB.183.14.4210-4216.2001 |
| [21] |
Winzer K, Hardie KR, Williams P. Bacterial cell-to-cell communication: sorry, can't talk now - gone to lunch!. Curr Opin Microbiol, 2002, 5(2): 216-222. DOI:10.1016/S1369-5274(02)00304-1 |
| [22] |
Lee JH, Lee J. Indole as an intercellular signal in microbial communities. FEMS Microbiol Rev, 2010, 34(4): 426-444. DOI:10.1111/j.1574-6976.2009.00204.x |
| [23] |
Lee JH, Kim YG, Baek KH, et al. The multifaceted roles of the interspecies signalling molecule indole in Agrobacterium tumefaciens. Environ Microbiol, 2015, 17(4): 1234-1244. DOI:10.1111/1462-2920.12560 |
| [24] |
Di Martino P, Fursy R, Bret L, et al. Indole can act as an extracellular signal to regulate biofilm formation of Escherichia coli and other indole-producing bacteria. Can J Microbiol, 2003, 49(7): 443-449. DOI:10.1139/w03-056 |
| [25] |
Chant EL, Summers DK. Indole signalling contributes to the stable maintenance of Escherichia coli multicopy plasmids. Mol Microbiol, 2007, 63(1): 35-43. DOI:10.1111/j.1365-2958.2006.05481.x |
| [26] |
Lee J, Jayaraman A, Wood TK. Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol, 2007, 7: 42. DOI:10.1186/1471-2180-7-42 |
| [27] |
Gaimster H, Cama J, Hernández-Ainsa S, et al. The indole pulse: a new perspective on indole signalling in Escherichia coli. PLoS ONE, 2014, 9(4): e93168. DOI:10.1371/journal.pone.0093168 |
| [28] |
Hirakawa H, Inazumi Y, Masaki T, et al. Indole induces the expression of multidrug exporter genes in Escherichia coli. Mol Microbiol, 2005, 55(4): 1113-1126. |
| [29] |
Nishino K, Senda Y, Yamaguchi A. The AraC-family regulator GadX enhances multidrug resistance in Escherichia coli by activating expression of mdtEF multidrug efflux genes. J Infect Chemother, 2008, 14(1): 23-29. DOI:10.1007/s10156-007-0575-Y |
| [30] |
Rahmati S, Yang S, Davidson AL, et al. Control of the AcrAB multidrug efflux pump by quorum-sensing regulator SdiA. Mol Microbiol, 2002, 43(3): 677-685. DOI:10.1046/j.1365-2958.2002.02773.x |
| [31] |
Domka J, Lee J, Wood TK. YliH (BssR) and YceP (BssS) regulate Escherichia coli K-12 biofilm formation by influencing cell signaling. Appl Environ Microbiol, 2006, 72(4): 2449-2459. DOI:10.1128/AEM.72.4.2449-2459.2006 |
| [32] |
Zhang XS, García-Contreras R, Wood TK. YcfR (BhsA) influences Escherichia coli biofilm formation through stress response and surface hydrophobicity. J Bacteriol, 2007, 189(8): 3051-3062. DOI:10.1128/JB.01832-06 |
| [33] |
Kuczyńska-Wiśnik D, Matuszewska E, Laskowska E. Escherichia coli heat-shock proteins IbpA and IbpB affect biofilm formation by influencing the level of extracellular indole. Microbiology, 2010, 156(1): 148-157. |
| [34] |
Lai Y, Xu ZL, Yan AX. A novel regulatory circuit to control indole biosynthesis protects Escherichia coli from nitrosative damages during the anaerobic respiration of nitrate. Environ Microbiol, 2017, 19(2): 598-610. DOI:10.1111/1462-2920.13527 |
| [35] |
Liu J, Summers D. Indole at low concentration helps exponentially growing Escherichia coli survive at high temperature. PLoS ONE, 2017, 12(12): e0188853. DOI:10.1371/journal.pone.0188853 |
| [36] |
Li X, Yang Q, Dierckens K, et al. RpoS and indole signaling control the virulence of Vibrio anguillarum towards gnotobiotic sea bass (Dicentrarchus labrax) larvae. PLoS ONE, 2014, 9(10): e111801. DOI:10.1371/journal.pone.0111801 |
| [37] |
Zheng J, Yu JJ, Jia MQ, et al. Indole enhances the survival of Pantoea ananatis YJ76 in face of starvation conditions. J Basic Microbiol, 2017, 57(7): 633-639. DOI:10.1002/jobm.201700027 |
| [38] |
Hidalgo-Romano B, Gollihar J, Brown SA, et al. Indole inhibition of N-acylated homoserine lactone-mediated quorum signalling is widespread in Gram-negative bacteria. Microbiology, 2014, 160(11): 2464-2473. |
| [39] |
Chu WH, Zere TR, Weber MM, et al. Indole production promotes Escherichia coli mixed-culture growth with Pseudomonas aeruginosa by inhibiting quorum signaling. Appl Environ Microbiol, 2012, 78(2): 411-419. DOI:10.1128/AEM.06396-11 |
| [40] |
Kim J, Park W. Indole inhibits bacterial quorum sensing signal transmission by interfering with quorum sensing regulator folding. Microbiology, 2013, 159(12): 2616-2625. |
| [41] |
Wang Y, Li H, Cui XX, et al. A novel stress response mechanism, triggered by indole, involved in quorum quenching enzyme MomL and iron-sulfur cluster in Muricauda olearia Th120. Sci Rep, 2017, 7(1): 4252. DOI:10.1038/s41598-017-04606-8 |
| [42] |
Oh S, Go GW, Mylonakis E, et al. The bacterial signalling molecule indole attenuates the virulence of the fungal pathogen Candida albicans. J Appl Microbiol, 2012, 113(3): 622-628. DOI:10.1111/j.1365-2672.2012.05372.x |
| [43] |
Blair JMA, Cloeckaert A, Nishino K, et al. Alternative explanation for indole-induced antibiotic tolerance in Salmonella. Proc Natl Acad Sci USA, 2013, 110(48): E4569. DOI:10.1073/pnas.1318318110 |
| [44] |
Lee JH, Cho HS, Kim Y, et al. Indole and 7-benzyloxyindole attenuate the virulence of Staphylococcus aureus. Appl Microbiol Biotechnol, 2013, 97(10): 4543-4552. DOI:10.1007/s00253-012-4674-z |
| [45] |
Molina‐Santiago C, Daddaoua A, Fillet S, et al. Interspecies signalling: Pseudomonas putida efflux pump TtgGHI is activated by indole to increase antibiotic resistance. Environ Microbiol, 2014, 16(5): 1267-1281. DOI:10.1111/1462-2920.12368 |
| [46] |
Vega NM, Allison KR, Samuels AN, et al. Salmonella typhimurium intercepts Escherichia coli signaling to enhance antibiotic tolerance. Proc Natl Acad Sci USA, 2013, 110(35): 14420-14425. DOI:10.1073/pnas.1308085110 |
| [47] |
Han Y, Yang CL, Yang Q, et al. Mutation of tryptophanase gene tnaA in Edwardsiella tarda reduces lipopolysaccharide production, antibiotic resistance and virulence. Environ Microbiol Rep, 2011, 3(5): 603-612. DOI:10.1111/j.1758-2229.2011.00269.x |
| [48] |
Monds RD, O'Toole GA. Metabolites as intercellular signals for regulation of community-level traits//Winans S, Bassler B, Eds. Chemical Communication among Bacteria. Washington, DC: ASM Press, 2008: 105-129.
|
| [49] |
Zarkan A, Cano-Muñiz S, Zhu JB, et al. Indole pulse signalling regulates the cytoplasmic pH of E. coli in a memory-like manner. Sci Rep, 2019, 9(1): 3868. DOI:10.1038/s41598-019-40560-3 |
| [50] |
Hirakawa H, Kodama T, Takumi-Kobayashi A, et al. Secreted indole serves as a signal for expression of type Ⅲ secretion system translocators in enterohaemorrhagic Escherichia coli O157:H7. Microbiology, 2009, 155(2): 541-550. |
| [51] |
Vega NM, Allison KR, Khalil AS, et al. Signaling-mediated bacterial persister formation. Nat Chem Biol, 2012, 8(5): 431-433. DOI:10.1038/nchembio.915 |
| [52] |
Kwan BW, Osbourne DO, Hu Y, et al. Phosphodiesterase DosP increases persistence by reducing cAMP which reduces the signal indole. Biotechnol Bioeng, 2015, 112(3): 588-600. DOI:10.1002/bit.25456 |
| [53] |
Vannini A, Volpari C, Gargioli C, et al. The crystal structure of the quorum sensing protein TraR bound to its autoinducer and target DNA. EMBO J, 2002, 21(17): 4393-4401. DOI:10.1093/emboj/cdf459 |
| [54] |
Bansal T, Englert D, Lee J, et al. Differential effects of epinephrine, norepinephrine, and indole on Escherichia coli O157:H7 chemotaxis, colonization, and gene expression. Infect Immun, 2007, 75(9): 4597-4607. DOI:10.1128/IAI.00630-07 |
| [55] |
Bansal T, Jesudhasan P, Pillai S, et al. Temporal regulation of enterohemorrhagic Escherichia coli virulence mediated by autoinducer-2. Appl Microbiol Biotechnol, 2008, 78(5): 811-819. DOI:10.1007/s00253-008-1359-8 |
| [56] |
Lee J, Zhang XS, Hegde M, et al. Indole cell signaling occurs primarily at low temperatures in Escherichia coli. ISME J, 2008, 2(10): 1007-1023. DOI:10.1038/ismej.2008.54 |
| [57] |
Maddela NR, Sheng BB, Yuan SS, et al. Roles of quorum sensing in biological wastewater treatment: a critical review. Chemosphere, 2019, 221: 616-629. DOI:10.1016/j.chemosphere.2019.01.064 |
| [58] |
Erb M, Veyrat N, Robert CA, et al. Indole is an essential herbivore-induced volatile priming signal in maize. Nat Commun, 2015, 6: 6273. DOI:10.1038/ncomms7273 |
| [59] |
Shen QQ, Liu LJ, Wang LP, et al. Indole primes plant defense against necrotrophic fungal pathogen infection. PLoS ONE, 2018, 13(11): e0207607. DOI:10.1371/journal.pone.0207607 |
| [60] |
Yu SM, Lee YH. Plant growth promoting rhizobacterium Proteus vulgaris JBLS202 stimulates the seedling growth of Chinese cabbage through indole emission. Plant Soil, 2013, 370(1/2): 485-495. |
| [61] |
Bhattacharyya D, Garladinne M, Lee YH. Volatile indole produced by rhizobacterium Proteus vulgaris JBLS202 stimulates growth of Arabidopsis thaliana through auxin, cytokinin, and brassinosteroid pathways. J Plant Growth Regul, 2015, 34(1): 158-168. DOI:10.1007/s00344-014-9453-x |
| [62] |
Yang Y, Gao KX, Wu Y, et al. Indole-3-acetic acid-mediated cross-kingdom signalling involved in plant-bacteria interactions. Biotechnol Bull, 2016, 32(8): 14-21 (in Chinese). 杨扬, 高克祥, 吴岩, 等. 吲哚乙酸跨界信号调节植物与细菌互作. 生物技术通报, 2016, 32(8): 14-21. |
| [63] |
Bansal T, Alaniz RC, Wood TK, et al. The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc Natl Acad Sci USA, 2010, 107(1): 228-233. DOI:10.1073/pnas.0906112107 |
| [64] |
Jaglin M, Rhimi M, Philippe C, et al. Indole, a signaling molecule produced by the gut microbiota, negatively impacts emotional behaviors in rats. Front Neurosci, 2018, 12: 216. |
| [65] |
Chimerel C, Emery E, Summers DK, et al. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep, 2014, 9(4): 1202-1208. DOI:10.1016/j.celrep.2014.10.032 |
| [66] |
Hubbard TD, Murray IA, Bisson WH, et al. Adaptation of the human aryl hydrocarbon receptor to sense microbiota-derived indoles. Sci Rep, 2015, 5: 12689. DOI:10.1038/srep12689 |
| [67] |
Hamaue N, Yamazaki N, Terado M, et al. Urinary isatin concentrations in patients with Parkinson's disease determined by a newly developed HPLC-UV method. Res Commun Mol Pathol Pharmacol, 2000, 108(1/2): 63-73. |
| [68] |
Moroni F, Carpenedo R, Venturini I, et al. Oxindole in pathogenesis of hepatic encephalopathy. Lancet, 1998, 351(9119): 1861. |
| [69] |
Zhao ZH, Lai JKL, Qiao L, et al. Role of gut microbial metabolites in nonalcoholic fatty liver disease. J Dig Dis, 2019, 20(4): 181-188. DOI:10.1111/1751-2980.12709 |
| [70] |
De Mello VD, Paananen J, Lindström J, et al. Indolepropionic acid and novel lipid metabolites are associated with a lower risk of type 2 diabetes in the Finnish Diabetes Prevention Study. Sci Rep, 2017, 7: 46337. DOI:10.1038/srep46337 |
| [71] |
Rajasekharan SK, Lee JH, Ravichandran V, et al. Assessments of iodoindoles and abamectin as inducers of methuosis in pinewood nematode, Bursaphelenchus xylophilus. Sci Rep, 2017, 7(1): 6803. DOI:10.1038/s41598-017-07074-2 |
| [72] |
Lee JH, Kim YG, Kim M, et al. Indole-associated predator-prey interactions between the nematode Caenorhabditis elegans and bacteria. Environ Microbiol, 2017, 19(5): 1776-1790. DOI:10.1111/1462-2920.13649 |
| [73] |
Kim J, Park W. Indole: a signaling molecule or a mere metabolic byproduct that alters bacterial physiology at a high concentration?. J Microbiol, 2015, 53(7): 421-428. DOI:10.1007/s12275-015-5273-3 |