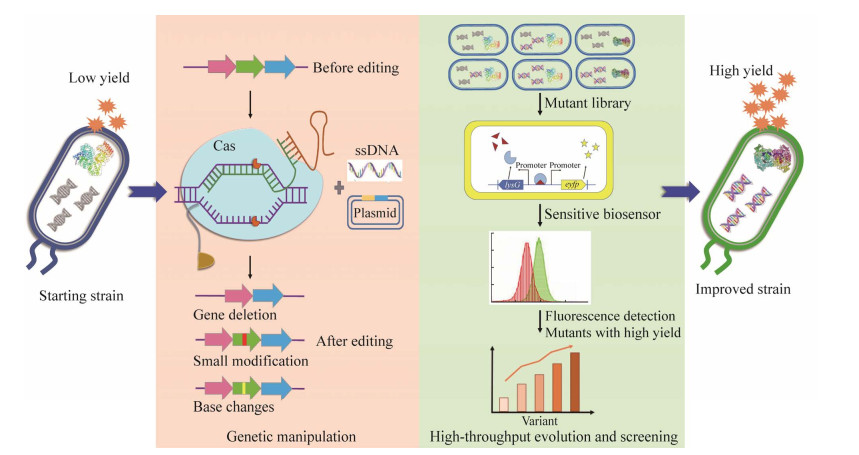
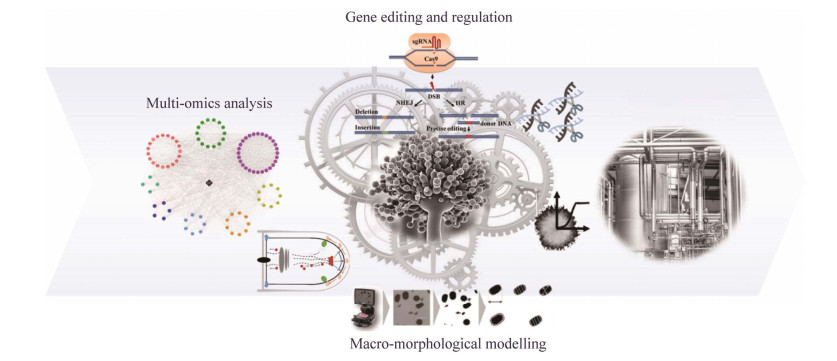
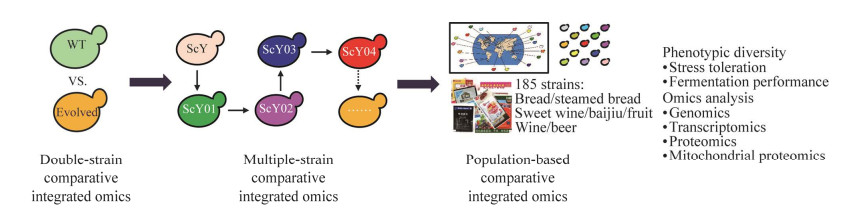
| [1] |
Ikeda M, Nakagawa S. The Corynebacterium glutamicum genome: features and impacts on biotechnological processes. Appl Microbiol Biotechnol, 2003, 62: 99-109. DOI:10.1007/s00253-003-1328-1
|
|
| [2] |
Zhao NN, Qian L, Luo GJ, et al. Synthetic biology approaches to access renewable carbon source utilization in Corynebacterium glutamicum. Appl Microbiol Biotechnol, 2018, 102(22): 9517-9529. DOI:10.1007/s00253-018-9358-x
|
|
| [3] |
Becker J, Rohles CM, Wittmann C. Metabolically engineered Corynebacterium glutamicum for bio-based production of chemicals, fuels, materials, and healthcare products. Metab Eng, 2018, 50: 122-141. DOI:10.1016/j.ymben.2018.07.008
|
|
| [4] |
Ruan YL, Zhu LJ, Li Q. Improving the electro- transformation efficiency of Corynebacterium glutamicum by weakening its cell wall and increasing the cytoplasmic membrane fluidity. Biotechnol Lett, 2015, 37(12): 2445-2452. DOI:10.1007/s10529-015-1934-x
|
|
| [5] |
Liu J, Wang Y, Lu YJ, et al. Mutations in peptidoglycan synthesis gene ponA improve electrotransformation efficiency of Corynebacterium glutamicum ATCC 13869. Appl Environ Microbiol, 2018, 84(24): e02225-18.
|
|
| [6] |
Jiang Y, Qian FH, Yang JJ, et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat Commun, 2017, 8: 15179. DOI:10.1038/ncomms15179
|
|
| [7] |
Zhang J, Qian FH, Dong F, et al. De novo engineering of Corynebacterium glutamicum for L-proline production. ACS Synth Biol, 2020, 9(7): 1897-1906. DOI:10.1021/acssynbio.0c00249
|
|
| [8] |
Krumbach K, Sonntag CK, Eggeling L, et al. CRISPR/Cas12a mediated genome editing to introduce amino acid substitutions into the mechanosensitive channel MscCG of Corynebacterium glutamicum. ACS Synth Biol, 2019, 8(12): 2726-2734. DOI:10.1021/acssynbio.9b00361
|
|
| [9] |
Liu J, Wang Y, Lu YJ, et al. Development of a CRISPR/Cas9 genome editing toolbox for Corynebacterium glutamicum. Microb Cell Fact, 2017, 16(1): 205. DOI:10.1186/s12934-017-0815-5
|
|
| [10] |
Liu J, Liu MS, Shi T, et al. CRISPR-assisted rational flux-tuning and arrayed CRISPRi screening of an L-proline exporter for L-proline hyperproduction. Nat Commun, 2022, 13: 891. DOI:10.1038/s41467-022-28501-7
|
|
| [11] |
Wang Y, Liu Y, Liu J, et al. MACBETH: multiplex automated Corynebacterium glutamicum base editing method. Metab Eng, 2018, 47: 200-210. DOI:10.1016/j.ymben.2018.02.016
|
|
| [12] |
Wang Y, Liu Y, Li JW, et al. Expanding targeting scope, editing window, and base transition capability of base editing in Corynebacterium glutamicum. Biotechnol Bioeng, 2019, 116(11): 3016-3029. DOI:10.1002/bit.27121
|
|
| [13] |
Deng C, Lv XQ, Li JH, et al. Development of a DNA double-strand break-free base editing tool in Corynebacterium glutamicum for genome editing and metabolic engineering. Metab Eng Commun, 2020, 11: e00135. DOI:10.1016/j.mec.2020.e00135
|
|
| [14] |
Wang Y, Cheng HJ, Liu Y, et al. In-situ generation of large numbers of genetic combinations for metabolic reprogramming via CRISPR-guided base editing. Nat Commun, 2021, 12: 678. DOI:10.1038/s41467-021-21003-y
|
|
| [15] |
Mustafi N, Grünberger A, Kohlheyer D, et al. The development and application of a single-cell biosensor for the detection of L-methionine and branched-chain amino acids. Metab Eng, 2012, 14(4): 449-457. DOI:10.1016/j.ymben.2012.02.002
|
|
| [16] |
Mustafi N, Grünberger A, Mahr R, et al. Application of a genetically encoded biosensor for live cell imaging of L-valine production in pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum strains. PLoS One, 2014, 9(1): e85731. DOI:10.1371/journal.pone.0085731
|
|
| [17] |
Binder S, Schendzielorz G, Stäbler N, et al. A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level. Genome Biol, 2012, 13(5): R40. DOI:10.1186/gb-2012-13-5-r40
|
|
| [18] |
Schendzielorz G, Dippong M, Grünberger A, et al. Taking control over control: use of product sensing in single cells to remove flux control at key enzymes in biosynthesis pathways. ACS Synth Biol, 2014, 3(1): 21-29. DOI:10.1021/sb400059y
|
|
| [19] |
Maike K, Christina M, Meike B, et al. Pyruvate carboxylase variants enabling improved lysine production from glucose identified by biosensor-based high-throughput fluorescence-activated cell sorting screening. ACS Synth Biol, 2019, 8(2): 274-281. DOI:10.1021/acssynbio.8b00510
|
|
| [20] |
Zheng B, Ma XY, Wang N, et al. Utilization of rare codon-rich markers for screening amino acid overproducers. Nat Commun, 2018, 9: 3616. DOI:10.1038/s41467-018-05830-0
|
|
| [21] |
Sun X, Li QG, Wang Y, et al. Isoleucyl-tRNA synthetase mutant based whole-cell biosensor for high-throughput selection of isoleucine overproducers. Biosens Bioelectron, 2021, 172: 112783. DOI:10.1016/j.bios.2020.112783
|
|
| [22] |
Chen JZ, Wang Y, Zheng P, et al. Engineering synthetic auxotrophs for growth-coupled directed protein evolution. Trends Biotechnol, 2022, 40(7): 773-776. DOI:10.1016/j.tibtech.2022.01.010
|
|
| [23] |
Dele-Osibanjo T, Li QJ, Zhang XL, et al. Growth-coupled evolution of phosphoketolase to improve L-glutamate production by Corynebacterium glutamicum. Appl Microbiol Biotechnol, 2019, 103(20): 8413-8425. DOI:10.1007/s00253-019-10043-6
|
|
| [24] |
Fu G, Yue J, Li DD, et al. An operator-based expression toolkit for Bacillus subtilis enables fine-tuning of gene expression and biosynthetic pathway regulation. PNAS, 2022, 119(11): e2119980119. DOI:10.1073/pnas.2119980119
|
|
| [25] |
Yu WW, Jin K, Wu YK, et al. A pathway independent multi-modular ordered control system based on thermosensors and CRISPRi improves bioproduction in Bacillus subtilis. Nucleic Acids Res, 2022, 50(11): 6587-6600. DOI:10.1093/nar/gkac476
|
|
| [26] |
Xu KD, Tong Y, Li Y, et al. Efficient, flexible autoinduction expression systems with broad initiation in Bacillus subtilis. ACS Synth Biol, 2021, 10(11): 3084-3093. DOI:10.1021/acssynbio.1c00369
|
|
| [27] |
Ye J, Li YJ, Bai YQ, et al. A facile and robust T7-promoter-based high-expression of heterologous proteins in Bacillus subtilis. Bioresour Bioprocess, 2022, 9(1): 1-12. DOI:10.1186/s40643-021-00489-w
|
|
| [28] |
丁明珠, 李炳志, 王颖, 等. 合成生物学重要研究方向进展. 合成生物学, 2020, 1(1): 7-28. Ding MZ, Li BZ, Wang Y, et al. Significant research progress in synthetic biology. Synth Biol J, 2020, 1(1): 7-28 (in Chinese).
|
|
| [29] |
Fang H, Li D, Kang J, et al. Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B 12. Nat Commun, 2018, 9: 4917. DOI:10.1038/s41467-018-07412-6
|
|
| [30] |
Jiang PT, Fang H, Zhao J, et al. Optimization of hydrogenobyrinic acid biosynthesis in Escherichia coli using multi-level metabolic engineering strategies. Microb Cell Fact, 2020, 19(1): 118. DOI:10.1186/s12934-020-01377-2
|
|
| [31] |
Li D, Fang H, Gai YM, et al. Metabolic engineering and optimization of the fermentation medium for vitamin B 12 production in Escherichia coli. Bioprocess Biosyst Eng, 2020, 43(10): 1735-1745. DOI:10.1007/s00449-020-02355-z
|
|
| [32] |
Tong ZY, Zheng XM, Tong Y, et al. Systems metabolic engineering for citric acid production by Aspergillus niger in the post-genomic era. Microb Cell Fact, 2019, 18(1): 28. DOI:10.1186/s12934-019-1064-6
|
|
| [33] |
Meyer V, Fiedler M, Nitsche B, et al. The cell factory Aspergillus enters the big data era: opportunities and challenges for optimising product formation. Adv Biochem Eng Biotechnol, 2015, 149: 91-132.
|
|
| [34] | |
|
| [35] | |
|
| [36] |
Kuivanen J, Arvas M, Richard P. Clustered genes encoding 2-keto-L-gulonate reductase and L-idonate 5-dehydrogenase in the novel fungal D-glucuronic acid pathway. Front Microbiol, 2017, 8: 225.
|
|
| [37] |
Kuivanen J, Wang YMJ, Richard P. Engineering Aspergillus niger for galactaric acid production: elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Microb Cell Fact, 2016, 15(1): 1-9. DOI:10.1186/s12934-015-0402-6
|
|
| [38] |
Pohl C, Kiel JAKW, Driessen AJM, et al. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synth Biol, 2016, 5(7): 754-764. DOI:10.1021/acssynbio.6b00082
|
|
| [39] |
Nødvig CS, Nielsen JB, Kogle ME, et al. A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS One, 2015, 10(7): e0133085. DOI:10.1371/journal.pone.0133085
|
|
| [40] |
Zheng XM, Zheng P, Sun JB, et al. Heterologous and endogenous U6 snRNA promoters enable CRISPR/Cas9 mediated genome editing in Aspergillus niger. Fungal Biol Biotechnol, 2018, 5: 2. DOI:10.1186/s40694-018-0047-4
|
|
| [41] |
Zheng XM, Zheng P, Zhang K, et al. 5S rRNA promoter for guide RNA expression enabled highly efficient CRISPR/Cas9 genome editing in Aspergillus niger. ACS Synth Biol, 2019, 8(7): 1568-1574. DOI:10.1021/acssynbio.7b00456
|
|
| [42] |
Punt PJ, Dingemanse MA, Kuyvenhoven A, et al. Functional elements in the promoter region of the Aspergillus nidulans gpdA gene encoding glyceraldehyde-3-phosphate dehydrogenase. Gene, 1990, 93(1): 101-109. DOI:10.1016/0378-1119(90)90142-E
|
|
| [43] |
Ganzlin M, Rinas U. In-depth analysis of the Aspergillus niger glucoamylase ( glaA) promoter performance using high-throughput screening and controlled bioreactor cultivation techniques. J Biotechnol, 2008, 135(3): 266-271. DOI:10.1016/j.jbiotec.2008.04.005
|
|
| [44] |
Blumhoff M, Steiger MG, Marx H, et al. Six novel constitutive promoters for metabolic engineering of Aspergillus niger. Appl Microbiol Biotechnol, 2013, 97(1): 259-267. DOI:10.1007/s00253-012-4207-9
|
|
| [45] |
Lu YD, Zheng XM, Wang Y, et al. Evaluation of Aspergillus niger six constitutive strong promoters by fluorescent-auxotrophic selection coupled with flow cytometry: a case for citric acid production. J Fungi, 2022, 8(6): 568. DOI:10.3390/jof8060568
|
|
| [46] |
Zhang LH, Zheng XM, Cairns TC, et al. Disruption or reduced expression of the orotidine-5′-decarboxylase gene pyrG increases citric acid production: a new discovery during recyclable genome editing in Aspergillus niger. Microb Cell Fact, 2020, 19(1): 76. DOI:10.1186/s12934-020-01334-z
|
|
| [47] |
Meyer V, Wanka F, Van Gent J, et al. Fungal gene expression on demand: an inducible, tunable, and metabolism-independent expression system for Aspergillus niger. Appl Environ Microbiol, 2011, 77(9): 2975-2983. DOI:10.1128/AEM.02740-10
|
|
| [48] |
Pel HJ, De Winde JH, Archer DB, et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat Biotechnol, 2007, 25: 221-231. DOI:10.1038/nbt1282
|
|
| [49] |
Sun JB, Lu X, Rinas U, et al. Metabolic peculiarities of Aspergillus niger disclosed by comparative metabolic genomics. Genome Biol, 2007, 8(9): R182. DOI:10.1186/gb-2007-8-9-r182
|
|
| [50] |
Schäpe P, Kwon MJ, Baumann B, et al. Updating genome annotation for the microbial cell factory Aspergillus niger using gene co-expression networks. Nucleic Acids Res, 2019, 47(2): 559-569. DOI:10.1093/nar/gky1183
|
|
| [51] |
Zhang QQ, Zheng XM, Wang Y, et al. Comprehensive optimization of the metabolomic methodology for metabolite profiling of Corynebacterium glutamicum. Appl Microbiol Biotechnol, 2018, 102(16): 7113-7121. DOI:10.1007/s00253-018-9095-1
|
|
| [52] |
Zheng XM, Yu JD, Cairns TC, et al. Comprehensive improvement of sample preparation methodologies facilitates dynamic metabolomics of Aspergillus niger. Biotechnol J, 2019, 14(3): 1800315. DOI:10.1002/biot.201800315
|
|
| [53] |
Cairns TC, Zheng XM, Zheng P, et al. Moulding the mould: understanding and reprogramming filamentous fungal growth and morphogenesis for next generation cell factories. Biotechnol Biofuels, 2019, 12: 77. DOI:10.1186/s13068-019-1400-4
|
|
| [54] |
Cairns TC, Zheng XM, Zheng P, et al. Turning inside out: filamentous fungal secretion and its applications in biotechnology, agriculture, and the clinic. J Fungi (Basel), 2021, 7(7): 535. DOI:10.3390/jof7070535
|
|
| [55] |
Cairns TC, Feurstein C, Zheng XM, et al. A quantitative image analysis pipeline for the characterization of filamentous fungal morphologies as a tool to uncover targets for morphology engineering: a case study using aplD in Aspergillus niger. Biotechnol Biofuels, 2019, 12: 149. DOI:10.1186/s13068-019-1473-0
|
|
| [56] |
Zheng XM, Cairns TC, Ni XM, et al. Comprehensively dissecting the hub regulation of PkaC on high-productivity and pellet macromorphology in citric acid producing Aspergillus niger. Microb Biotechnol, 2022, 15(6): 1867-1882. DOI:10.1111/1751-7915.14020
|
|
| [57] |
Cairns TC, Feurstein C, Zheng XM, et al. Functional exploration of co-expression networks identifies a nexus for modulating protein and citric acid titres in Aspergillus niger submerged culture. Fungal Biol Biotechnol, 2019, 6: 18. DOI:10.1186/s40694-019-0081-x
|
|
| [58] |
Cairns TC, Zheng XM, Feurstein C, et al. A library of Aspergillus niger chassis strains for morphology engineering connects strain fitness and filamentous growth with submerged macromorphology. Front Bioeng Biotechnol, 2022, 9: 820088. DOI:10.3389/fbioe.2021.820088
|
|
| [59] |
齐显尼, 甘雨满, 王钦宏. 耐高温酿酒酵母的选育及其在乙醇发酵生产中的应用. 生物产业技术, 2018(4): 84-89. Qi XN, Gan YM, Wang QH. Development of thermotolerant Saccharomyces cerevisiae and its application in ethanol fermentation at high temperature. Biotechnol Bus, 2018(4): 84-89 (in Chinese).
|
|
| [60] |
Yao Z, Wang QH, Dai ZJ. Recent advances in directed yeast genome evolution. J Fungi (Basel), 2022, 8(6): 635. DOI:10.3390/jof8060635
|
|
| [61] |
Fraczek MG, Naseeb S, Delneri D. History of genome editing in yeast. Yeast, 2018, 35(5): 361-368. DOI:10.1002/yea.3308
|
|
| [62] |
Christian M, Cermak T, Doyle EL, et al. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics, 2010, 186(2): 757-761. DOI:10.1534/genetics.110.120717
|
|
| [63] |
DiCarlo JE, Norville JE, Mali P, et al. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res, 2013, 41(7): 4336-4343. DOI:10.1093/nar/gkt135
|
|
| [64] |
Zhang GQ, Lin YP, Qi XN, et al. TALENs-assisted multiplex editing for accelerated genome evolution to improve yeast phenotypes. ACS Synth Biol, 2015, 4(10): 1101-1111. DOI:10.1021/acssynbio.5b00074
|
|
| [65] |
Gan YM, Lin YP, Guo YF, et al. Metabolic and genomic characterisation of stress-tolerant industrial Saccharomyces cerevisiae strains from TALENs-assisted multiplex editing. FEMS Yeast Res, 2018, 18(5): foy045.
|
|
| [66] |
Vervoort Y, Linares AG, Roncoroni M, et al. High-throughput system-wide engineering and screening for microbial biotechnology. Curr Opin Biotechnol, 2017, 46: 120-125. DOI:10.1016/j.copbio.2017.02.011
|
|
| [67] | |
|
| [68] |
Wang Z, Lin YP, Dai ZJ, et al. Modulating DNA repair pathways to diversify genomic alterations in Saccharomyces cerevisiae. Microbiol Spectr, 2022, 10(2): e0232621. DOI:10.1128/spectrum.02326-21
|
|
| [69] |
Tan J, Forner J, Karcher D, et al. DNA base editing in nuclear and organellar genomes. Trends Genet, 2022, S0168-9525(22): 00176-7.
|
|
| [70] |
Liu YF, Lin YP, Guo YF, et al. Stress tolerance enhancement via SPT15 base editing in Saccharomyces cerevisiae. Biotechnol Biofuels, 2021, 14(1): 155. DOI:10.1186/s13068-021-02005-w
|
|
| [71] |
Shui WQ, Xiong Y, Xiao WD, et al. Understanding the mechanism of thermotolerance distinct from heat shock response through proteomic analysis of industrial strains of Saccharomyces cerevisiae. Mol Cell Proteomics, 2015, 14(7): 1885-1897. DOI:10.1074/mcp.M114.045781
|
|
| [72] |
Xiao WD, Duan XX, Lin YP, et al. Distinct proteome remodeling of industrial Saccharomyces cerevisiae in response to prolonged thermal stress or transient heat shock. J Proteome Res, 2018, 17(5): 1812-1825. DOI:10.1021/acs.jproteome.7b00842
|
|
| [73] |
Xiong Y, Guo YF, Xiao WD, et al. An NGS-independent strategy for proteome-wide identification of single amino acid polymorphisms by mass spectrometry. Anal Chem, 2016, 88(5): 2784-2791. DOI:10.1021/acs.analchem.5b04417
|
|
| [74] |
Gan YM, Qi XN, Lin YP, et al. A hierarchical transcriptional regulatory network required for long-term thermal stress tolerance in an industrial Saccharomyces cerevisiae strain. Front Bioeng Biotechnol, 2022, 9: 826238. DOI:10.3389/fbioe.2021.826238
|
|
| [75] |
Wang Z, Qi Q, Lin YP, et al. QTL analysis reveals genomic variants linked to high-temperature fermentation performance in the industrial yeast. Biotechnol Biofuels, 2019, 12: 59. DOI:10.1186/s13068-019-1398-7
|
|
| [76] |
Dzialo MC, Park R, Steensels J, et al. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol Rev, 2017, 41(Supp_1): S95-S128. DOI:10.1093/femsre/fux031
|
|
| [77] |
Liu GD, Chen Y, Færgeman NJ, et al. Elimination of the last reactions in ergosterol biosynthesis alters the resistance of Saccharomyces cerevisiae to multiple stresses. FEMS Yeast Res, 2017, 17(6): fox063.
|
|
| [78] |
Becker J, Wittmann C. From systems biology to metabolically engineered cells—an omics perspective on the development of industrial microbes. Curr Opin Microbiol, 2018, 45: 180-188. DOI:10.1016/j.mib.2018.06.001
|
|
 2022, Vol. 38
2022, Vol. 38