 2022, Vol. 38
2022, Vol. 38中国科学院微生物研究所、中国微生物学会主办
文章信息
- 李道明, 王瑛, 陈超, 曾明白, 李倩如, 贾青云, 刘秀丽, 侯勇跃, 范成明, 陈宇红, 胡赞民
- LI Daoming, WANG Ying, CHEN Chao, ZENG Mingbai, LI Qianru, JIA Qingyun, LIU Xiuli, HOU Yongyue, FAN Chengming, CHEN Yuhong, HU Zanmin
- 芽孢杆菌几种重要抗菌脂肽研究进展
- Advances in several important antimicrobial lipopeptids from Bacillus spp.
- 生物工程学报, 2022, 38(5): 1768-1783
- Chinese Journal of Biotechnology, 2022, 38(5): 1768-1783
- 10.13345/j.cjb.220001
-
文章历史
- Received: January 1, 2022
- Accepted: March 22, 2022
- Published: March 25, 2022

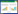
引用本文

|



2. 中国科学院遗传与发育生物学研究所 中国科学院种子创新研究院, 北京 100101;
3. 内蒙古农牧业科学院兽医研究所, 内蒙古 呼和浩特 010031;
4. 中国科学院大学 现代农业科学学院, 北京 101408
2. The Innovative Academy of Seed Design, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China;
3. Veterinary Research Institute, Inner Mongolia Academy of Agriculture and Animal Husbandry Sciences, Hohhot 010031, Inner Mongolia, China;
4. College of Advanced Agriculture Sciences, University of Chinese Academy of Sciences, Beijing 101408, China
芽孢杆菌属(Bacillus spp.) 细菌广泛地存在于生物体和环境中,是一类在农业、工业及医药卫生等领域具有重要应用价值的细菌[1],其中枯草芽孢杆菌(Bacillus subtilis) 是一种公认的益生菌,已广泛用于植保和动物饲料添加剂[2-3]。芽孢杆菌属细菌是防治植物病害的常规农药的最有前途的替代品,能够促进植物生长[4],诱导植物自身抗性[5],抑制不同病害,以及通过竞争空间和营养成分直接对微生物病原体造成拮抗作用[6]。虽然芽孢杆菌的有益保护作用涉及多种机制,但抗菌脂肽化合物的产生是影响其生防活性的最重要因素之一,因此抗菌脂肽的深入研究具有重要的理论意义和应用价值。
多数芽孢杆菌可产生低分子量、两亲性的脂肽。脂肽类化合物的结构和功能多样,性能稳定,表面活性优异,可广泛应用于农业、石油化工、环保、食品工业和生物医药等领域[7-11]。表面活性素(surfactin)、伊枯草菌素(iturin) 和芬原素(fengycin) 是脂肽的3个重要家族。这3个家族都是环脂肽,都含有一个不同C链长度的β-羟基脂肪酸链及由不同氨基酸组成的环状肽。由于C链长度不同及氨基酸组成不同,被分成不同的家族,具有不同的功能。前人对抗菌脂肽surfactin、iturin、fengycin在结构与功能、合成机理和调控等方面已取得重要研究进展。3种环脂肽结构明显不同,功能和应用价值有较大差异。研究显示surfactin不仅对细菌的运动和生物膜的形成意义非凡[12],它们的抗病毒/抗菌活性也使得表面活性素具有广泛用途[13]。fengycin和iturin通常以它们优越的抗真菌能力为人所熟知。三者都是通过非核糖体肽链合成酶合成的,但三者的合成基因及调控机制不同。目前通过合成基因调控元件的改造,能够使surfactin的产量大幅提高至12.8 g/L[14],为功能脂肽的规模生产开辟了新途径。本领域的研究进展虽然已有少量综述[8],但尚缺综合性进展总结和评述。本文将全面总结surfactin、iturin和fengycin的结构、功能、合成机理和调控、分离纯化和发酵生产以及用途等方面的研究进展,并对未来发展方向做了讨论,有助于更好地理解这三类脂肽产生的分子调控机制、作用机理、相关关系及潜在产品的研发和应用。
1 抗菌脂肽的结构和功能脂肽是一类由亲水的环状或短线性寡肽和疏水的长链脂肪酸两部分组成的结构高度多样性的生物表面活性剂,基于氨基酸结构的不同可分为环状脂肽和线性脂肽两类。脂肽结构的多样性除由氨基酸种类不同导致外,脂肪酸链的长度和分支差异也是主要原因。目前,26个家族中约90种脂肽化合物已被鉴定,其中24个家族中86种脂肽为环脂肽,只有剩下的2个家族4种脂肽为线性脂肽[15]。产脂肽的微生物主要是芽孢杆菌属细菌,如枯草芽孢杆菌(B. subtilis)、地衣芽孢杆菌(Bacillus licheniformis)、多粘芽孢杆菌(Bacillus polymyxa)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、甲基营养型芽孢杆菌(Bacillus methylotrophicus)、萎缩芽孢杆菌(Bacillus atrophaeus)、副(类) 地衣芽孢杆菌(Bacillus paralicheniformis)、短小芽孢杆菌(Bacillus pumilus)、蜡状芽孢杆菌(Bacillus cereus)、巨大芽孢杆菌(Bacillus megaterium)等。表面活性素(surfactin)、伊枯草菌素(iturin)、芬原素(fengycin)、杆菌霉素(bacillomycin)、多粘菌素(polymyxins) 等是芽孢杆菌中研究最广泛的脂肽[16]。单个菌株可以产生至少一种环脂肽,有的菌株可同时产生surfactin、iturin和fengycin[17-18]。脂肽具有广谱抑菌活性,在抑制真菌生长方面表现优异[19]。
1.1 SurfactinSurfactin是枯草芽孢杆菌产生的一类最主要的抗菌活性物质。1968年Arima等最初是将其作为一种有效的凝血抑制剂,并命名为surfactin[20]。从生物化学角度来看,surfactin包含17%的脂质和13%的肽链。典型的surfactin是由7个α-氨基酸残基和一个长度为C13−C15的羟基脂肪酸,以内酯键结合形成的环状化合物分子[21]。在水溶液中,surfactin带负电荷的Glu和Asp形成一个微小的极性结构域,在折叠的骨架中肽环呈“马鞍”状拓扑结构,促成各种各样的生物学特性[22]。surfactin在肽环中典型的氨基酸顺序是:l-Glu1-l-Leu2-d-Leu3-l- Val4-l-Asp5-d-leu6-l-Leu7[23],如图 1A所示。由于肽链上第2、4、7位氨基酸不保守,脂肪酸中碳原子数不同及构型的差异导致surfactin具有众多同系物和同分异构体。1991年Baumgart等报道了在典型肽链组成的第7位l-Leu分别被l-Val和l-Ile所取代的两种surfactin同系物,surfactin A (Val7) 和surfactin B (Ile7)[24]。1994年Peypoux等又报道了一种在典型肽链组成的第4位l-Val被l-Ala所取代的surfactin (Ala4)[25],随后的研究更精确地测定了十余种同系物。不同亚型的surfactin具有细微的物理化学特性差异。

|
| 图 1 Surfactin、iturin和fengycin的一级结构[28-29] Fig. 1 Primary structures of three lipopeptids. (A) surfactin, n=9–11. (B) iturin, n=14–17. (C) fengycin, n=11–15. n: the number of CH2 groups in the peptide chain. l and d are l-amino acids and d-amino acids in the α-amino acid residues, respectively. Modified from [28-29]. |
| |
通常,脂肽的杀菌活性随着脂肪酸链长度(通常为C10–C12) 的增加而增强,脂肪酸链含有较多碳原子(14或16) 的脂肽具有更强抗细菌活性或抗真菌活性[26]。surfactin碳链的长度影响其灭活病毒的能力,随着脂肪酸链碳原子数目增多,疏水性增加,灭活病毒的能力增强[27]。
Surfactin具有独特的两亲结构,是优秀的生物表面活性剂,能够降低表面和界面张力,具有良好的起泡、增溶、润湿、去垢和乳化特性,在工业中具有广泛的应用价值[30]。其中对原油良好的乳化作用,可应用于石油污染处理和高效、可持续石油回收,强化石油开采,降解柴油[31]。surfactin还能有效减小海水和油之间的张力,改变环境的润湿指数,因此具有截留油的能力[32]。
Surfactin具有较强抗菌作用。Krishnan等在体外发现surfactin对串珠镰刀菌(Fusarium moniliforme) 抑菌效果比多菌灵更显著[33]。Park等发现贝莱斯芽孢杆菌(Bacillus velezensis) GH1-13分泌的surfactin对多种植物病原真菌有较强的拮抗作用,明显抑制菌丝生长[34]。
除此之外surfactin还表现出抗病毒、抗肿瘤和抗支原体活性,具有非常强的溶血功能[35-37]。Qi等报道Bacillus amyloliquefaciens WH1产生一种表面活性素,命名为WH1 fungin[38]。Gao等发现在以小鼠为模型的实验中WH1 fungin能够改善Ⅰ型糖尿病[39]。
生物膜是一种结构复杂的多细胞微生物群落,细胞外多聚物能够保护细胞,并牢固黏在物质表面。surfactin在去除病原体形成的生物膜方面表现出优异的性能。许多研究证明,surfactin能够降低细菌粘附作用[40]。低浓度的surfactin与单一细菌或混合病原体短时间接触就能破坏其生物膜。因此,在去除病原微生物在外科手术用具上形成的生物膜,保护生物材料方面,surfactin抗细菌黏着性和抗细菌形成生物膜特性具有潜在的医学应用价值。
值得注意的是,有研究发现surfactin还具有潜在蚊虫杀蛹作用。Geetha等分离的一株芽孢杆菌分泌的surfactin对埃及伊蚊(Aedes aegypti)、埃及按蚊(Anopheles stephensi) 和埃及库蚊(Culex quinquefasciatus) 具有灭蚊活性[41],可用于防治病媒传播疾病等公共防控项目。
1.2 IturinIturin是一大类强烈抑制真菌生长的脂肽类化合物。在结构上与surfactin较为类似,同样包含一个由7个α-氨基酸残基组成的肽链,是一种环脂七肽。七肽的第7位氨基酸是Ser,由14–17个C原子的脂肪酸链的β-氨基酸残基连接(图 1B)。七肽的氨基酸序列为(l-)Asn-(d-)Tyr-(d-)Asn-(l-)Gln-(l-)Pro-(d-)Asn- (l-)Ser。该家族主要包括的亚型有:iturin A、C、D、E,杆菌抗霉素(bacillomycin) D、F、L、Lc和抗霉枯草菌素(mycosubtilin)、杆菌肽素(bacillopeptin) 等[42]。其中最具有代表性的iturin A由一个含有23–17个碳原子的β-氨基脂肪酸和7个氨基酸残基组成的环状结构,其环状结构由7位的Ser的羧基和β-氨基脂肪酸的氨基缩合形成[43]。Ser6-Asn7互换便成为mycosubtilin,它的抗菌活性比iturin更高。而bacillomycin的基本结构和iturin A类似,只是在1、4、5、6和7位上的氨基酸有所不同,分为bacillomycin D、F、L、Lc。
Iturin对黄瓜(Cucumis sativus) 细菌性叶斑病和软腐病、小麦(Triticum aestivum) 赤霉病有着很好的防控作用[44]。此外iturin有很强的溶血作用,但其生理活性和surfactin不同,对真菌(包括酵母菌) 具有强烈的拮抗作用,但是对细菌拮抗作用很小[45]。研究表明,伊枯草菌素对红色毛癣菌具有较强抑制作用[46]。Iturins抗菌谱广,稳定性高,毒性低,致敏性低,应用前景广。
1.3 FengycinFengycin由β-羟基脂肪酸和10个氨基酸残基的小肽组成,经典结构如图 1C所示。脂肪酸链往往由14−18个碳原子组成,肽链包含4个D-氨基酸和6个l-氨基酸,肽链中氨基酸组成顺序为l-Glu1-d-Orn2-l-Tyr3-d-Thr4-l-Glu5- d-Ala6 (Val)6-l-Pro7-l-Gln8-d-Tyr9-l-Ile10,肽链的第10位上的Ile羧基和第3位的Tyr上羟基缩合形成环状结构[47]。
Fengycin主要有两种类型(fengycin A和fengycin B),当肽链的第6位上是Ala时,属于fengycin A,而当肽链的第6位上是Val时,属于fengycin B[48],分子量在1 500 Da左右。制磷脂菌素(plipstatin) 是由B. cereus BMG302-fF67和B. subtilis 168产生的一种和fengycin非常类似的菌脂肽,其结构及性质和fengycin大致相同,目前多数文献将其归类于fengycin系列[49]。
Fengycin具有良好的抑真菌活性,尤其对丝状真菌更为明显。对细菌和酵母无作用[50],能抑制芳香酶和磷脂酶A2的活性。此外,fengycin还具有溶解红细胞作用[51]。Piewngam等证明枯草芽孢杆菌产生的fengycin能抑制金黄色葡萄球菌(Staphylococcus aureus) 在小鼠体内定殖[52]。
1.4 Surfactin、iturin及fengycin结构和功能的比较芽孢杆菌产生的surfactin、iturin及fengycin都是环脂肽类小分子活性物质,都含有亲水性肽链和亲脂族烃链,具有双亲性,都具有多种生物活性,但三者一级结构明显不同,功能也有显著差异,因此其应用领域也有不同(表 1)。脂肽具有多种生物活性(如抗菌、抗病毒、溶血和杀虫等),脂肽一个很重要的特性是使生物膜穿孔,即通过直接作用于细菌和真菌的生物膜,使膜形成孔隙并导致渗透性不平衡,从而使细胞死亡。这种特性可以帮助脂肽的生物活性得以实现[53]。脂肽或分泌脂肽的菌剂已被尝试作为抑制各种植物病原体的多功能武器,对包括细菌、真菌和卵菌在内的多种植物病原体均有拮抗作用。Iturin和fengycin具有抗真菌活性,而surfactin具有更广泛的强效抗菌活性,也被用作幼虫杀虫剂[54-55]。
| Items | Surfactin | Iturin | Fengycin | References |
| Molecular weight | About 1 036 Da | About 1 057 Da | About 1 463 Da | [21, 43, 47] |
| The length of fatty acid chain | β-hydroxy fatty acid chain, C13–C16 | β-hydroxy fatty acid chain, C14–C17 | β-hydroxy fatty acid chain, C14–C18 | [21, 42, 47] |
| The length of cyclic peptide chain | Seven α-amino acids | Seven α-amino acids | Ten α-amino acids | [21, 42, 47] |
| l-Glu1-l-Leu2-d-Leu3-l-Val4-l-Asp5-d-Leu6-l-Leu7 | l-Asn1-d-Tyr2-d-Asn3-l- Gln4-l-Pro5-d-Asn6-l-Ser7 | l-Glu1-d-Orn2-l-Tyr3-d- Thr4-l-Glu5-d-Ala6 (Val)6- l-Pro7-l-Gln8-d-lyr9-l-Ile10 | [21, 42, 47] | |
| Subtypes | Surfactin A, Surfactin B Esperin, Halobacillin Pumilacidin |
Iturin A, Iturin C, Iturin D Iturin E, Bacillomycin D Bacillomycin F, Bacillomycin L, Bacillomycin Lc, Mycosubtilin, Bacillopeptin |
Fengycin A Fengycin B Plipastatin A Plipastatin B |
[53] |
| Main activity | Strong surfactant, antifungal, antibacterial, antiviral, anti-tumor, anti-mycoplasma, biofilm removal, strong hemolysis and pore formation | Surfactant and antifungal | Surfactant and antifungal | [16, 53] |
| Applications | Biopesticides (plant disease control, such as sclerotinia, stem rot, and soybean root and leaf diseases); Medicine (antiviral, anti-fungal, anti-bacterial, anti-cancer, anti-thrombotic); Cosmetics (emulsifiers, antibacterial agents); Food safety (biological surfactants and emulsifiers); and oil extraction, etc. | Biopesticides (plant disease control, such as sclerotinia, stem rot, soybean blight, and powdery mildew of melon) | Biopesticides (plant disease control, such as soybean blight, sclerotinia, stem rot, apple gray mold, and melon powdery mildew etc.) | [16, 54-55] |
抗菌脂肽的合成不同于传统的以mRNA为模板、tRNA为载体转录翻译合成蛋白质的模式,是通过非核糖体肽链合成酶(nonribosomal peptide synthetase, NRPS) 合成的[56-57]。NRPS是一种能识别、激活、修饰和连接氨基酸中间体和产物肽的多模块酶,是目前所发现的最大酶系。它由起始、伸长和终止3个主要模块形成脂肽合成的组装线,每一个模块由不同的结构域构成(图 2A)。它的底物可以是标准的氨基酸或甲基化、羟基化、氯化或其他修饰的变体,但也可能不同于氨基酸如β-或环丙基-氨基酸或α-羟基酸。NRPS首先通过腺苷酰化结构域(adenylation domain, A domain) 识别、结合底物,并将由ATP衍生的AMP添加到底物上使其激活;激活的底物与4′-磷酸泛酰巯基乙胺中游离巯基结合,且4′-磷酸泛酰巯基乙胺绑定在紧邻A domain下游的肽酰基载体蛋白结构域(peptide carrier protein, PCP-domain,也常称之为T domain) 上;在缩合结构域(condensation domain, C domain) 的作用下与前一个模块的肽链相连接,该过程按照相邻合成酶各组成模块的顺序依次向前直到多肽合成结束。非核糖体肽合成酶亚基的最后一个模板中通常包含一个差向异构酶结构(epimerization domain, E domain),其作用是将被激活的l-氨基酸转化为d-氨基酸,某些合成酶模块也能直接活化D构象的氨基酸[58-59]。综上所述,全酶可以由多个模块按照特定的空间顺序排列而成,模块的数量、种类和排列顺序决定肽链的长短、氨基酸的种类和顺序。非核糖体肽合成机理如图 2所示。

|
| 图 2 非核糖体肽合成(NRPS) 的多模块模型[61] Fig. 2 Multi-module model of non-ribosomal peptide synthesis (NRPS) (Modified from [61]). A schematic arrangement of domains within a hypothetical three module NRPS that contains an initiation (blue), an elongation (red) and a termination (green) module. A domain: adenylation domain; T domain: aminoacyl carrier domain; C domain: condensation domain; TE: thioester domain; It is in turn responsible for the activation and transfer of amino acid substrates, the formation of peptide bonds, the termination of extension and the release of products on the domain. |
| |
2019年Reimer等在非核糖体肽合成酶的功能方面取得重要进展,该团队采用X射线晶体技术,为NRPSs拍摄了超高分辨率的3D图像,第一次看到NRPSs的各个模块,对单个模块与更大的组装线之间的关系进行细致的观察。这项研究发现,除了模块必须协调才能将中间体从一个工作站传递到下一个工作站外,其他所有模块之间都缺乏同步性,各个模块不是以直线或其他有组织的方式排列,而是可以在许多不同的相对位置排列,NRPSs具有高度的灵活性。该研究透彻解析了脂肽化合物合成的分子机制,人们可尝试在基因水平上重新设计非核糖体肽合成酶每个模块或结构域的排列方式,通过混合和匹配模块来设计NRPSs,从而得到具有新的氨基酸组成的脂肽分子,为获得新的抗菌药物提供基础[60]。
目前已有通过合成模块的删除、截短改良抗菌脂肽的范例。Gao等删除fengycin合成酶复合体单个模块使整个酶复合体失活,但删除第7个模块的A/T domain产生了截短的线性脂肽(C16–C17β-OHFA-Glu-Orn-Tyr-Thr-Glu-Ala/ Val),删除A6 domain发生模块跳跃现象,产生3个新型的fengycin亚型,五肽(C16–17β- OHFA-Glu-Orn-Tyr-Thr-Glu)、六肽(C16–17β- OHFA-Glu-Orn-Tyr-Thr-Glu-Ile) 和八肽(C16–17β- OHFA-Glu-Orn-Tyr-Thr-Glu-Gln-Tyr-Ile)[62],但对新型脂肽的生物活性并没有相关研究。
2.1 Surfactin的生物合成及其调控Surfactin合成酶基因srfAA、srfAB、srfAC、srfAD共同构成srfA操纵子,基因总长达27 kb,由srfAA和srfAB编码蛋白负责组装前6位氨基酸,srfAC编码蛋白组装第7位氨基酸。srfAC编码的第一个硫酯结构域TE负责终止肽链延伸并释放多肽产物,srfAD编码的第二个硫酯酰基转移结构域(thioesterase/acyltransferase domain, TE/AT domain) 则具有激活并启动surfactin肽链合成反应的功能[63]。此外,在srfA操纵子的下游,含有一个surfactin合成必需的磷酸泛酰巯基乙胺基转移酶(PPTases) 基因sfp,行使激活surfactin合成酶的功能[64]。枯草芽孢杆菌产生的脂肽类抗生素生物合成的基因调控研究最为详细的是surfactin。Surfactin合成的基因调控是由复杂的群感效应系统ComQXPA诱导的多重级联反应过程。芽孢杆菌信息素ComX感受细胞密度,随着细胞密度增加而增加,当细胞密度达到一定阈值,信息素ComX与膜蛋白组氨酸激酶ComP结合,诱导ComP的自磷酸化。磷酸化的ComP进而将磷酸基团转移至调控蛋白ComA。磷酸化的ComA与surfactin合成酶基因srfA的启动子特定区域结合,激活RNA聚合酶,使srfA开始转录[65-67]。由rap基因编码的天冬氨酰基磷酸酶与下游phr基因编码的感受态刺激因子CSF结合后,丧失磷酸酶活性,以致丧失对磷酸化ComA的脱磷酸化作用,使ComA处于活化状态,利于srfA基因的转录和surfactin的合成[68]。负调控因子还有Rap (天冬氨酸磷酸酶)、CodY (GTP和支链氨基酸依赖的转录调节因子)、PerR (Fur家族转录因子)、SinI (转录调控因子)、Spx (调节蛋白)、PhoP (双元件响应调节器)。其中CodY通过直接作用srfA启动子抑制其表达,Spx通过阻断在启动子区域ComA与RNA聚合酶作用从而抑制surfactin合成[69],SinI通过正向调节生物膜的形成直接调控surfactin合成[70]。全局调控因子DegU对surfactin合成起正调控作用。敲除degU基因,导致srfA基因的转录显著下降。此外,由基因swrC (即yreP) 和liaIHGFSR操纵子编码的蛋白负责surfactin转运和细胞对surfactin耐受性[71]。
多种枯草芽孢杆菌可生产surfactin,但产量较低(< 1 g/L)。Jiao等利用启动子改造,使重组菌株THY-7/Pg3 surfactin产量达到了9.74 g/L[72]。Wu等采用系统代谢工程方法构建surfactin的高产菌,最终surfactin产量增加至12.8 g/L,产率为65.0 mmol/mol蔗糖(理论产率的42%)[14]。这些研究为surfactin的规模生产提供了新的技术途径[73]。
2.2 Iturin的生物合成及其调控Iturin家族的脂肽类抗菌素受不同基因的调控。其中bacillomycin D由bam/bmy基因簇负责合成,mycosubtilin由myc基因簇负责合成,iturin A由itu基因簇编码负责合成。itu基因簇包括4个开放阅读框(ORFs),分别为ituA、ituB、ituC、和ituD[74-75]。其中bam/bmy基因簇分别发现存在于B. subtilis AU195和B. amyloliquefaciens FZB42中,该基因簇编码一个多功能复合酶,包括一个脂肪酸合成酶、一个氨基转移酶和一个肽合成酶。第一个开放阅读框架bmyD、ituD和fenF编码丙二酰辅酶A转移酶,第二个开放阅读框bmyA、ituA和mycA分别编码乙酰辅酶A连接酶、酰基载体蛋白(CAP)、β-酮乙基合成酶和转氨酶结构域。Iturin家族中mycosubtilin的基因调控首次在合成mycosubtilin的菌株B. subtilis ATCC 6633得到阐明,myc操纵子的表达依赖于ComA,与群体响感应有关。SigH因子和Spo0H也影响myc操纵子表达,培养基中添加PhrC蛋白可以弥补缺失spo0H缺失突变体产生的现象。AbrB可以抑制myc表达,敲除abrB基因可以增强myc表达[76]。类似于fen操纵子,基因degQ也是bmy操纵子的一个正向调控因子。类似于srfA操纵子,全局调控因子DegU和ComA对bmy操纵子的全转录激活必不可少,DegU直接作用于bmy操纵子启动子上游的两个位点,对bmy表达起重要作用。而且DegU和跨膜蛋白YczE对bacillomycin转录后调控起重要作用。与fengycin和surfactin等脂肽一样,类似于Sfp蛋白的Lpa-14对iturin的合成必不可少[77]。将强启动子C2up插入itu操纵子的上游,可提高iturins产量[78]。
2.3 Fengycin的生物合成及其调控Fengycin生物合成基因簇包含5个开放阅读框ORFs,分别编码fengycin合成酶的5个亚基,fen操纵子包含fenC (编码287 kDa蛋白)、fenD (编码290 kDa蛋白)、fenE (编码286 kDa蛋白)、fenA (编码406 kDa蛋白) 和fenB (编码146 kDa蛋白),它们线性排列共享一个启动子[79]。同样地,基因sfp对fengycin的合成必不可少,B. subtilis 168虽然含有完整的fen操纵子,但由于sfp中间含有一个终止密码子,导致Sfp蛋白不完整,不能合成fengycin[14]。但仅转化sfp基因的B. subtilis 168菌株fengycin产量仍然很低,原因是其degQ的启动子区–10位置上的一个碱基T突变成了C,因此该菌株不能表达DegQ[80]。DegQ是一种胞外蛋白酶增强子,编码一个含有46个氨基酸的多肽,是广泛存在于芽孢杆菌中的多效性因子,控制多种水解酶的产生。degQ基因的敲除会导致fengycin同系物合成水平明显下降[81]。研究还发现,通过对B. subtilis F29-3中fen操纵子的转录分析,RNA聚合酶可以结合在fen操纵子启动子上游富含A和T的被称为UP元件区域,从而增强fen操纵子的转录[82]。多核苷酸磷酸化酶对fengycins合成有重要影响,pnpA基因失活时fengycins产量大幅下降[83]。Spo0A蛋白和ComA蛋白通过与fen启动子结合等机制促进fengycins的产生[84]。近年来,fengycins的生产工艺也有了较大改进,如人工全固相合成脱羟基fengycins衍生物技术和细胞固定化生产fengycins技术[85-86],为规模生产fengycins奠定了基础。
3 脂肽发酵生产和分离纯化尽管抗菌活性脂肽具有多种应用潜力,但是其实际应用因野生型菌株产量低而受到较大限制。采用固定化细胞发酵[87]、泡沫回流发酵、无泡发酵[88]、固态发酵[89]、调节供氧水平和补充金属元素等手段对生产工艺和培养基成分进行优化,或采用分批发酵或补料分批发酵方式已成功使枯草芽孢杆菌合成抗菌脂肽物质的产量得到大幅度的提升。2006年Yeh等报道以ppch作为固定载体经过72 h的发酵,B. subtilis ATCC 21332菌株surfactin的产量达6.45 g/L,增加2.0−4.3倍[90];在LB为培养基发酵液中添加0.35%二价铁离子后,B. subtilis ATCC 21332菌株fengycin产量由对照组的310 mg/L提高到680 mg/L,提高了近1.1倍[86]。
在脂肽研究中,脂肽物质的分离、提纯和鉴定是相当重要的一部分,菌体发酵液中含有多种未知的蛋白质等大分子物质和小分子活性物质,而且大多数微生物可产生两种以上的抑菌脂肽物质。在此混合体系中,抗菌脂肽物质的分离纯化程度需视研究工作的类型而定,至今还没有单独或一种现成的方法可以把任何一种脂肽物质从复杂的混合物中提取并纯化出来,往往通过几种方法联合使用。
目前,对于分析级的抗菌脂肽的分离纯化往往联合有机溶剂萃取,酸沉淀,硫酸铵沉淀、透析等两种以上的方法得到粗提物,再经过离子交换层析、固相萃取(SPE)、聚苯乙烯树脂等方法得到进一步纯化的抗菌脂肽(表 2)。不同技术有优缺点不同(表 2),对于制备级的抗菌脂肽的分离纯化常采用膜超滤技术、吸附法、泡沫分离法。
| Extraction/purification methods | Advantages | Disadvantages | References |
| Precipitation by acid treatment | It is an effective method for recovery of crude lipopeptides and can be applied on large scale. It cost low and operation process is simple |
The product is acidic | [91-94] |
| Precipitation by ammonium sulfate | The operation process is simple | Dialysis is required for reducing salt pollution | [95-96] |
| Extraction by organic solvent | It is an effective method for recovery of crude lipopeptides and can be applied on large scale. The organic solvent for the lipopeptide extraction can be reused | It is expensive and toxic | [30, 97-99] |
| Separation by collection of foam | The product possessed high purity | It required complicated equipment | [100-102] |
| Membrane ultrafiltration | The process is fast and the product possessed high purity | It is expensive and must be operated under high pressure | [103-104] |
| Ion exchange chromatography | The process is fast and the product possess high purity. Resin can be reused | Different effective resin materials need to be selected | [105-107] |
| Adsorption resin | The product possessed high purity and high quality. Resin can be reused. The process is fast | Desorption is need with organic solvent | [108] |
| High performance liquid chromatography (HPLC) | The product possessed high purity. It can be used for fractionation of different lipopeptides, the identification of lipopeptide isomers, the qualitative and quantitative analysis of lipopeptides | Retention rate is low | [108-109] |
| Thin layer chromatography (TLC) | It can be used for identification of cyclic lipopeptides | The retention rate is very low and is generally used to identify lipopeptides | [110-112] |
| Gel filtration | Matrix can be reused | The retention rate is very low | [98, 113-114] |
尽管芽孢杆菌产生的脂肽研究已取得了较大进展,并且具有应用价值,但脂肽作为商品在市场上的应用还很初步。迄今为止,仅有少部分国外商家能够小规模生产出商业化surfactin、iturin和fengycin等产品,如Sigma和OKA公司生产的脂肽产品。而国内还没有此类商业化产品,即便是国外生产的标准品,同一分子量的脂肽仍包含多种同分异构体。然而能够产生脂肽的菌剂已在国内外广泛应用,如用于饲料添加剂的枯草芽孢杆菌、用于医药的地衣芽孢杆菌等。
鉴于脂肽潜在的应用价值及目前研究存在的局限性,未来需要在下述方面加强研究。1)筛选天然高产菌株。2) 脂肽的安全性尚缺乏系统的研究。安全性研究是食品、药品及农业领域应用的先决条件,目前虽然有一些脂肽应用研究,但涉及其安全性研究有限。3) 高产廉价脂肽的生产技术。目前的脂肽生产主要是利用天然菌株或工程菌株探讨其生产工艺,产量不高。目前只有少量脂肽的合成生物学研究,以提高脂肽的产量。例如,Jiao等[72]、Wu等[14]将surfactin的产量分别提升至9.74 g/L和12.8 g/L。但还缺乏完善的规模生产、分离纯化工艺流程。需要开发更高效的生产工艺及构建更高产脂肽的工程菌。利用廉价的农业废弃物作为生产原料以降低生产成本也是未来研究的方向。4) 高活性脂肽研究。高活性脂肽是未来研究的重要目标,可通过合成生物学增加或减少脂肪酸链的长度或改变肽链中氨基酸成分获得活性改变的脂肽,目前缺乏更高活性的脂肽化合物。5) 具有新功能脂肽的获得。微生物可产生多种天然脂肽,不同微生物分泌的脂肽种类也不同,有些脂肽含量很低,但可能具有新的生物学功能,对微量新脂肽的鉴定比较困难,需要较高的分离纯化技术、结构分析技术。
5 结论脂肽是微生物分泌的重要化合物,具有抗菌、抗病毒、抗炎症、抗肿瘤等多种功能,其结构、功能及合成调控研究已取得重要进展,其应用潜力巨大,可用于种植业、养殖业、食品、医药、石油工业等领域。未来脂肽的发展需要在新型脂肽的发现、高产活性脂肽的生产、脂肽生产的低成本技术研发及合成脂肽的安全性等方面加强。
| [1] |
Wei XX, Xin D, Xin YH, et al. Bacillus depressus sp.nov., isolated from soil of a sunflower field. Antonie Van Leeuwenhoek, 2016, 109(1): 13-20. DOI:10.1007/s10482-015-0605-1
|
| [2] |
Du YP, Xu ZC, Yu GL, et al. A newly isolated Bacillus subtilis strain named WS-1 inhibited diarrhea and death caused by pathogenic Escherichia coli in newborn piglets. Front Microbiol, 2019, 10: 1248. DOI:10.3389/fmicb.2019.01248
|
| [3] |
Liu P, Zhao JB, Guo PT, et al. Dietary corn bran fermented by Bacillus subtilis MA139 decreased gut cellulolytic bacteria and microbiota diversity in finishing pigs. Front Cell Infect Microbiol, 2017, 7: 526. DOI:10.3389/fcimb.2017.00526
|
| [4] |
Verma SK, White JF. Indigenous endophytic seed bacteria promote seedling development and defend against fungal disease in browntop millet (Urochloa ramosa L.). J Appl Microbiol, 2018, 124: 764-778. DOI:10.1111/jam.13673
|
| [5] |
Kloepper JW, Ryu CM, Zhang S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathol, 2004, 94: 1259-1266. DOI:10.1094/PHYTO.2004.94.11.1259
|
| [6] |
Shafi J, Tian H, Ji M. Bacillus species as versatile weapons for plant pathogens: a review. Biotechnol Biotechnol Equip, 2017, 31: 446-459. DOI:10.1080/13102818.2017.1286950
|
| [7] |
Lucera A, Costa C, Conte A, et al. Food applications of natural antimicrobial compounds. Front Microbiol, 2012, 3: 287.
|
| [8] |
周伟, 郑维, 陶思美, 等. 芽胞杆菌产生的环脂肽类物质的研究进展. 微生物学杂志, 2015, 35(4): 80-86. Zhou W, Zheng W, Tao SM, et al. Advance in cyclic lipopeptide substances from Bacillus subtilis. J Microbiol, 2015, 35(4): 80-86 (in Chinese). DOI:10.3969/j.issn.1005-7021.2015.04.014 |
| [9] |
Sumi CD, Yang BW, Yeo IC, et al. Antimicrobial peptides of the genus Bacillus: a new era for antibiotics. Can J Microbiol, 2015, 61: 93-103. DOI:10.1139/cjm-2014-0613
|
| [10] |
Carolin FC, Kumar PS, Ngueagni PT. A review on new aspects of lipopeptide biosurfactant: types, production, properties and its application in the bioremediation process. J Hazard Mater, 2021, 407: 124827. DOI:10.1016/j.jhazmat.2020.124827
|
| [11] |
Zhu ZW, Zhang BY, Cai QH, et al. A critical review on the environmental application of lipopeptide micelles. Bioresour Technol, 2021, 339: 125602. DOI:10.1016/j.biortech.2021.125602
|
| [12] |
Zeriouh H, De Vicente A, Perez-Garcia A, et al. Surfactin triggers biofilm formation of Bacillus subtilis in melon phylloplane and contributes to the biocontrol activity. Environ Microbiol, 2014, 16: 2196-2211. DOI:10.1111/1462-2920.12271
|
| [13] |
Saggese A, Culurciello R, Casillo A, et al. A marine isolate of Bacillus pumilus secretes a pumilacidin active against Staphylococcus aureus. Mar Drugs, 2018, 16: 180. DOI:10.3390/md16060180
|
| [14] |
Wu Q, Zhi Y, Xu Y. Systematically engineering the biosynthesis of a green biosurfactant surfactin by Bacillus subtilis 168. Metab Eng, 2019, 52: 87-97. DOI:10.1016/j.ymben.2018.11.004
|
| [15] |
Liu JF, Mbadinga SM, Yang SZ, et al. Chemical structure, property and potential applications of biosurfactants produced by Bacillus subtilis in petroleum recovery and spill mitigation. Int J Mol Sci, 2015, 16(3): 4814-4837. DOI:10.3390/ijms16034814
|
| [16] |
Meena KR, Kanwar SS. Lipopeptides as the antifungal and antibacterial agents: applications in food safety and therapeutics. Biomed Res Int, 2015, 2015: 473050.
|
| [17] |
Kim PI, Ryu J, Kim YH, et al. Production of biosurfactant lipopeptides iturin A, fengycin and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J Microbiol Biotechnol, 2010, 20(1): 138-145. DOI:10.4014/jmb.0905.05007
|
| [18] |
Roongsawang N, Thaniyavarn J, Thaniyavarn S, et al. Isolation and characterization of a halotolerant Bacillus subtilis BBK-1 which produces three kinds of lipopeptides: bacillomycin L, plipastatin, and surfactin. Extremophiles, 2002, 6(6): 499-506. DOI:10.1007/s00792-002-0287-2
|
| [19] |
Hamley IW. Lipopeptides: from self-assembly to bioactivity. Chem Commun (Camb), 2015, 51(41): 8574-8583. DOI:10.1039/C5CC01535A
|
| [20] |
ARIMA K, Kakinuma A, Tamura G. Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: isolation, characterization and its inhibition of fibrin clot formation. Biochem Biophys Res Commun, 1968, 31(3): 488-494. DOI:10.1016/0006-291X(68)90503-2
|
| [21] |
Shao CS, Liu L, Gang HZ, et al. Structural diversity of the microbial surfactin derivatives from selective esterification approach. Int J Mol Sci, 2015, 16(1): 1855-1872. DOI:10.3390/ijms16011855
|
| [22] |
Jin H, Li KP, Niu YX, et al. Continuous enhancement of iturin A production by Bacillus subtilis with a stepwise two-stage glucose feeding strategy. BMC Biotechnol, 2015, 15: 53. DOI:10.1186/s12896-015-0172-6
|
| [23] |
Liu S, Wang H, Zhang J, et al. In vitro mutation and selection of doubled-haploid Brassica napus lines with improved resistance to Sclerotinia sclerotiorum. Plant Cell Rep, 2005, 24(3): 133-144. DOI:10.1007/s00299-005-0925-0
|
| [24] |
Baumgart F, Kluge B, Ullrich C, et al. Identification of amino acid substitutions in the lipopeptide surfactin using 2D NMR spectroscopy. Biochem Biophys Res Commun, 1991, 177(3): 998-1005. DOI:10.1016/0006-291X(91)90637-M
|
| [25] |
Peypoux F, Bonmatin JM, Labbe H, et al. [Ala4] surfactin, a novel isoform from Bacillus subtilis studied by mass and NMR spectroscopies. Eur J Biochem, 1994, 224(1): 89-96. DOI:10.1111/j.1432-1033.1994.tb19998.x
|
| [26] |
Baindara P, Mandal SM, Chawla N, et al. Characterization of two antimicrobial peptides produced by a halotolerant Bacillus subtilis strain SK.DU. 4 isolated from a rhizosphere soil sample. AMB Express, 2013, 3(1): 2. DOI:10.1186/2191-0855-3-2
|
| [27] |
Kracht M, Rokos H, Ozel M, et al. Antiviral and hemolytic activities of surfactin isoforms and their methyl ester derivatives. J Antibiot (Tokyo), 1999, 52(7): 613-619. DOI:10.7164/antibiotics.52.613
|
| [28] |
Yang H, Li X, Li X, et al. Identification of lipopeptide isoforms by MALDI-TOF-MS/MS based on the simultaneous purification of iturin, fengycin, and surfactin by RP-HPLC. Anal Bioanal Chem, 2015, 407(9): 2529-2542. DOI:10.1007/s00216-015-8486-8
|
| [29] |
Zhao HB, Shao DY, Jiang CM, et al. Biological activity of lipopeptides from Bacillus. Appl Microbiol Biotechnol, 2017, 101(15): 5951-5960. DOI:10.1007/s00253-017-8396-0
|
| [30] |
Kim KM, Lee JY, Kim CK, et al. Isolation and characterization of surfactin produced by Bacillus polyfermenticus KJS-2. Arch Pharm Res, 2009, 32(5): 711-715. DOI:10.1007/s12272-009-1509-2
|
| [31] |
Al-Wahaibi Y, Joshi S, Al-Bahry S, et al. Biosurfactant production by Bacillus subtilis B30 and its application in enhancing oil recovery. Colloids Surf B Biointerfaces, 2014, 114: 324-333. DOI:10.1016/j.colsurfb.2013.09.022
|
| [32] |
Youssef N, Simpson DR, Duncan KE, et al. In situ biosurfactant production by Bacillus strains injected into a limestone petroleum reservoir. Appl Environ Microbiol, 2007, 73(4): 1239-1247. DOI:10.1128/AEM.02264-06
|
| [33] |
Krishnan N, Velramar B, Velu RK. Investigation of antifungal activity of surfactin against mycotoxigenic phytopathogenic fungus Fusarium moniliforme and its impact in seed germination and mycotoxicosis. Pestic Biochem Physiol, 2019, 155: 101-107. DOI:10.1016/j.pestbp.2019.01.010
|
| [34] |
Park G, Nam J, Kim J, et al. Structure and mechanism of surfactin peptide from Bacillus velezensis antagonistic to fungi plant pathogens. Bull Korean Chem Soc, 2019, 40(7): 704-709. DOI:10.1002/bkcs.11757
|
| [35] |
Deravel J, Lemière S, Coutte F, et al. Mycosubtilin and surfactin are efficient, low ecotoxicity molecules for the biocontrol of lettuce downy mildew. Appl Microbiol Biotechnol, 2014, 98(14): 6255-6264. DOI:10.1007/s00253-014-5663-1
|
| [36] |
Duarte C, Gudiña EJ, Lima CF, et al. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express, 2014, 4: 40. DOI:10.1186/s13568-014-0040-0
|
| [37] |
Liu XY, Tao XY, Zou AH, et al. Effect of the microbial lipopeptide on tumor cell lines: apoptosis induced by disturbing the fatty acid composition of cell membrane. Protein Cell, 2010, 1(6): 584-594. DOI:10.1007/s13238-010-0072-4
|
| [38] |
Qi GF, Zhu FY, Du P, et al. Lipopeptide induces apoptosis in fungal cells by a mitochondria-dependent pathway. Peptides, 2010, 31(11): 1978-1986. DOI:10.1016/j.peptides.2010.08.003
|
| [39] |
Gao ZQ, Zhao XY, Yang T, et al. Immunomodulation therapy of diabetes by oral administration of a surfactin lipopeptide in NOD mice. Vaccine, 2014, 32(50): 6812-6819. DOI:10.1016/j.vaccine.2014.08.082
|
| [40] |
Loiseau C, Schlusselhuber M, Bigot R, et al. Surfactin from Bacillus subtilis displays an unexpected anti-Legionella activity. Appl Microbiol Biotechnol, 2015, 99(12): 5083-5093. DOI:10.1007/s00253-014-6317-z
|
| [41] |
Geetha I, Manonmani AM. Surfactin: a novel mosquitocidal biosurfactant produced by Bacillus subtilis ssp.subtilis (VCRC B471) and influence of abiotic factors on its pupicidal efficacy. Lett Appl Microbiol, 2010, 51(4): 406-412. DOI:10.1111/j.1472-765X.2010.02912.x
|
| [42] |
Ali S, Hameed S, Imran A, et al. Genetic, physiological and biochemical characterization of Bacillus sp.strain RMB7 exhibiting plant growth promoting and broad spectrum antifungal activities. Microb Cell Fact, 2014, 13: 144.
|
| [43] |
Hiradate S, Yoshida S, Sugie H, et al. Mulberry anthracnose antagonists (iturins) produced by Bacillus amyloliquefaciens RC-2. Phytochemistry, 2002, 61(6): 693-698. DOI:10.1016/S0031-9422(02)00365-5
|
| [44] |
Gong AD, Li HP, Yuan QS, et al. Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS One, 2015, 10(2): e0116871. DOI:10.1371/journal.pone.0116871
|
| [45] |
Klich MA, Lax AR, Bland JM. Inhibition of some mycotoxigenic fungi by iturin A, a peptidolipid produced by Bacillus subtilis. Mycopathologia, 1991, 116(2): 77-80. DOI:10.1007/BF00436368
|
| [46] |
Cotta SR, Da Mota FF, Tupinambá G, et al. Antimicrobial activity of Paenibacillus kribbensis POC 115 against the dermatophyte Trichophyton rubrum. World J Microbiol Biotechnol, 2012, 28(3): 953-962. DOI:10.1007/s11274-011-0893-1
|
| [47] |
Cochrane SA, Vederas JC. Lipopeptides from Bacillus and Paenibacillus spp.: a gold mine of antibiotic candidates. Med Res Rev, 2016, 36(1): 4-31. DOI:10.1002/med.21321
|
| [48] |
Tang QY, Bie XM, Lu ZX, et al. Effects of fengycin from Bacillus subtilis fmbJ on apoptosis and necrosis in Rhizopus stolonifer. J Microbiol, 2014, 52(8): 675-680. DOI:10.1007/s12275-014-3605-3
|
| [49] |
Tsuge K, Ano T, Hirai M, et al. The genes degQ, pps, and lpa-8 (sfp) are responsible for conversion of Bacillus subtilis 168 to plipastatin production. Antimicrob Agents Chemother, 1999, 43(9): 2183-2192. DOI:10.1128/AAC.43.9.2183
|
| [50] |
Vanittanakom N, Loeffler W, Koch U, et al. Fengycin—a novel antifungal lipopeptide antibiotic produced by Bacillus subtilis F-29-3. J Antibiot (Tokyo), 1986, 39(7): 888-901. DOI:10.7164/antibiotics.39.888
|
| [51] |
Cheng W, Feng YQ, Ren J, et al. Anti-tumor role of Bacillus subtilis fmbJ-derived fengycin on human colon cancer HT29 cell line. Neoplasma, 2016, 63(2): 215-222.
|
| [52] |
Piewngam P, Zheng Y, Nguyen TH, et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature, 2018, 562(7728): 532-537. DOI:10.1038/s41586-018-0616-y
|
| [53] |
Falardeau J, Wise C, Novitsky L, et al. Ecological and mechanistic insights into the direct and indirect antimicrobial properties of Bacillus subtilis lipopeptides on plant pathogens. J Chem Ecol, 2013, 39(7): 869-878. DOI:10.1007/s10886-013-0319-7
|
| [54] |
Cossus L, Roux-Dalvai F, Kelly I, et al. Interactions with plant pathogens influence lipopeptides production and antimicrobial activity of Bacillus subtilis strain PTB185. Biol Control, 2021, 154: 104497. DOI:10.1016/j.biocontrol.2020.104497
|
| [55] |
Maksimov IV, Singh BP, Cherepanova EA, et al. Prospects and applications of lipopeptide-producing bacteria for plant protection (review). Appl Biochem Microbiol, 2020, 56(1): 15-28. DOI:10.1134/S0003683820010135
|
| [56] |
Doekel S, Marahiel MA. Biosynthesis of natural products on modular peptide synthetases. Metab Eng, 2001, 3(1): 64-77. DOI:10.1006/mben.2000.0170
|
| [57] |
Marahiel MA. Protein templates for the biosynthesis of peptide antibiotics. Chem Biol, 1997, 4(8): 561-567. DOI:10.1016/S1074-5521(97)90242-8
|
| [58] |
Wu XC, Qian CD, Fang HH, et al. Paenimacrolidin, a novel macrolide antibiotic from Paenibacillus sp.F6-B70 active against methicillin-resistant Staphylococcus aureus. Microb Biotechnol, 2011, 4(4): 491-502. DOI:10.1111/j.1751-7915.2010.00201.x
|
| [59] |
Marahiel MA. A structural model for multimodular NRPS assembly lines. Nat Prod Rep, 2016, 33(2): 136-140. DOI:10.1039/C5NP00082C
|
| [60] |
Reimer JM, Eivaskhani M, Harb I, et al. Structures of a dimodular nonribosomal peptide synthetase reveal conformational flexibility. Science, 2019, 366(6466): eaaw4388. DOI:10.1126/science.aaw4388
|
| [61] |
Calcott MJ, Ackerley DF. Genetic manipulation of non-ribosomal peptide synthetases to generate novel bioactive peptide products. Biotechnol Lett, 2014, 36(12): 2407-2416. DOI:10.1007/s10529-014-1642-y
|
| [62] |
Gao L, Guo JP, Fan Y, et al. Module and individual domain deletions of NRPS to produce plipastatin derivatives in Bacillus subtilis. Microb Cell Fact, 2018, 17(1): 84. DOI:10.1186/s12934-018-0929-4
|
| [63] |
Peypoux F, Bonmatin JM, Wallach J. Recent trends in the biochemistry of surfactin. Appl Microbiol Biotechnol, 1999, 51(5): 553-563. DOI:10.1007/s002530051432
|
| [64] |
Mootz HD, Finking R, Marahiel MA. 4′-phosphopantetheine transfer in primary and secondary metabolism of Bacillus subtilis. J Biol Chem, 2001, 276(40): 37289-37298. DOI:10.1074/jbc.M103556200
|
| [65] |
Comella N, Grossman AD. Conservation of genes and processes controlled by the quorum response in bacteria: characterization of genes controlled by the quorum-sensing transcription factor ComA in Bacillus subtilis. Mol Microbiol, 2005, 57(4): 1159-1174. DOI:10.1111/j.1365-2958.2005.04749.x
|
| [66] |
Oslizlo A, Stefanic P, Dogsa I, et al. Private link between signal and response in Bacillus subtilis quorum sensing. PNAS, 2014, 111(4): 1586-1591. DOI:10.1073/pnas.1316283111
|
| [67] |
Tran LS, Nagai T, Itoh Y. Divergent structure of the ComQXPA quorum-sensing components: molecular basis of strain-specific communication mechanism in Bacillus subtilis. Mol Microbiol, 2000, 37(5): 1159-1171. DOI:10.1046/j.1365-2958.2000.02069.x
|
| [68] |
Auchtung JM, Lee CA, Grossman AD. Modulation of the ComA-dependent quorum response in Bacillus subtilis by multiple Rap proteins and Phr peptides. J Bacteriol, 2006, 188: 5273-5285. DOI:10.1128/JB.00300-06
|
| [69] |
Zhang Y, Nakano S, Choi SY, et al. Mutational analysis of the Bacillus subtilis RNA polymerase α C-terminal domain supports the interference model of spx-dependent repression. J Bacteriol, 2006, 188(12): 4300-4311. DOI:10.1128/JB.00220-06
|
| [70] |
López D, Vlamakis H, Losick R, et al. Paracrine signaling in a bacterium. Genes Dev, 2009, 23(14): 1631-1638. DOI:10.1101/gad.1813709
|
| [71] |
Li X, Yang H, Zhang DL, et al. Overexpression of specific proton motive force-dependent transporters facilitate the export of surfactin in Bacillus subtilis. J Ind Microbiol Biotechnol, 2015, 42(1): 93-103. DOI:10.1007/s10295-014-1527-z
|
| [72] |
Jiao S, Li X, Yu HM, et al. In situ enhancement of surfactin biosynthesis in Bacillus subtilis using novel artificial inducible promoters. Biotechnol Bioeng, 2017, 114(4): 832-842. DOI:10.1002/bit.26197
|
| [73] |
梅雨薇, 杨子云, 于樊, 等. 生物表面活性剂脂肽的发酵生产及抑菌应用研究进展. 中国生物工程杂志, 2020, 40(5): 105-116. Mei YW, Yang ZY, Yu F, et al. Recent progress on fermentation and antibacterial applications of surfactin. China Biotechnol, 2020, 40(5): 105-116 (in Chinese). |
| [74] |
Moyne AL, Cleveland TE, Tuzun S. Molecular characterization and analysis of the operon encoding the antifungal lipopeptide bacillomycin D. FEMS Microbiol Lett, 2004, 234(1): 43-49. DOI:10.1111/j.1574-6968.2004.tb09511.x
|
| [75] |
Tsuge K, Akiyama T, Shoda M. Cloning, sequencing, and characterization of the iturin A operon. J Bacteriol, 2001, 183(21): 6265-6273. DOI:10.1128/JB.183.21.6265-6273.2001
|
| [76] |
Duitman EH, Wyczawski D, Boven LG, et al. Novel methods for genetic transformation of natural Bacillus subtilis isolates used to study the regulation of the mycosubtilin and surfactin synthetases. Appl Environ Microbiol, 2007, 73(11): 3490-3496. DOI:10.1128/AEM.02751-06
|
| [77] |
Huang CC, Ano T, Shoda M. Nucleotide sequence and characteristics of the gene, lpa-14, responsible for biosynthesis of the lipopeptide antibiotics iturin A and surfactin from Bacillus subtilis RB14. J Ferment Bioeng, 1993, 76(6): 445-450. DOI:10.1016/0922-338X(93)90238-4
|
| [78] |
Dang YL, Zhao FJ, Liu XS, et al. Enhanced production of antifungal lipopeptide iturin A by Bacillus amyloliquefaciens LL3 through metabolic engineering and culture conditions optimization. Microb Cell Fact, 2019, 18(1): 68. DOI:10.1186/s12934-019-1121-1
|
| [79] |
Wu CY, Chen CL, Lee YH, et al. Nonribosomal synthesis of fengycin on an enzyme complex formed by fengycin synthetases. J Biol Chem, 2007, 282(8): 5608-5616. DOI:10.1074/jbc.M609726200
|
| [80] |
Yang M, Ferrari E, Chen E, et al. Identification of the pleiotropic sacQ gene of Bacillus subtilis. J Bacteriol, 1986, 166(1): 113-119. DOI:10.1128/jb.166.1.113-119.1986
|
| [81] |
Wang PP, Guo QG, Ma YN, et al. DegQ regulates the production of fengycins and biofilm formation of the biocontrol agent Bacillus subtilis NCD-2. Microbiol Res, 2015, 178: 42-50. DOI:10.1016/j.micres.2015.06.006
|
| [82] |
Ke WJ, Chang BY, Lin TP, et al. Activation of the promoter of the fengycin synthetase operon by the UP element. J Bacteriol, 2009, 191(14): 4615-4623. DOI:10.1128/JB.00255-09
|
| [83] |
Yaseen Y, Diop A, Gancel F, et al. Polynucleotide phosphorylase is involved in the control of lipopeptide fengycin production in Bacillus subtilis. Arch Microbiol, 2018, 200(5): 783-791. DOI:10.1007/s00203-018-1483-5
|
| [84] |
Zhao JF, Zhang C, Lu ZX. Differential proteomics research of Bacillus amyloliquefaciens and its genome-shuffled saltant for improving fengycin production. Braz J Microbiol, 2018, 49(Suppl 1): 166-177. |
| [85] |
Rosés C, Camó C, Oliveras À, et al. Total solid-phase synthesis of dehydroxy fengycin derivatives. J Org Chem, 2018, 83(24): 15297-15311. DOI:10.1021/acs.joc.8b02553
|
| [86] |
Hmidet N, Ben Ayed H, Jacques P, et al. Enhancement of surfactin and fengycin production by Bacillus mojavensis A21: application for diesel biodegradation. Biomed Res Int, 2017, 2017: 5893123.
|
| [87] |
Gancel F, Montastruc L, Liu T, et al. Lipopeptide overproduction by cell immobilization on iron-enriched light polymer particles. Process Biochem, 2009, 44(9): 975-978. DOI:10.1016/j.procbio.2009.04.023
|
| [88] |
Keller K, Friedmann T, Boxman A. The bioseparation needs for tomorrow. Trends Biotechnol, 2001, 19(11): 438-441. DOI:10.1016/S0167-7799(01)01803-0
|
| [89] |
Willenbacher J, Rau JT, Rogalla J, et al. Foam-free production of surfactin via anaerobic fermentation of Bacillus subtilis DSM 10(T). AMB Express, 2015, 5: 21. DOI:10.1186/s13568-015-0107-6
|
| [90] |
Chtioui O, Dimitrov K, Gancel F, et al. Biosurfactants production by immobilized cells of Bacillus subtilis ATCC 21332 and their recovery by pertraction. Process Biochem, 2010, 45(11): 1795-1799. DOI:10.1016/j.procbio.2010.05.012
|
| [91] |
Thaniyavarn J, Roongsawang N, Kameyama T, et al. Production and characterization of biosurfactants from Bacillus licheniformis F2.2. Biosci Biotechnol Biochem, 2003, 67(6): 1239-1244. DOI:10.1271/bbb.67.1239
|
| [92] |
Wei YH, Wang LC, Chen WC, et al. Production and characterization of fengycin by indigenous Bacillus subtilis F29-3 originating from a potato farm. Int J Mol Sci, 2010, 11(11): 4526-4538. DOI:10.3390/ijms11114526
|
| [93] |
Chitarra GS, Breeuwer P, Nout MJR, et al. An antifungal compound produced by Bacillus subtilis YM 10-20 inhibits germination of Penicillium roqueforti conidiospores. J Appl Microbiol, 2003, 94(2): 159-166. DOI:10.1046/j.1365-2672.2003.01819.x
|
| [94] |
Gong M, Wang JD, Zhang J, et al. Study of the antifungal ability of Bacillus subtilis strain PY-1 in vitro and identification of its antifungal substance (iturin A). Acta Biochim Biophys Sin, 2006, 38(4): 233-240. DOI:10.1111/j.1745-7270.2006.00157.x
|
| [95] |
Moyne AL, Shelby R, Cleveland TE, et al. Bacillomycin D: an iturin with antifungal activity against Aspergillus flavus. J Appl Microbiol, 2001, 90(4): 622-629. DOI:10.1046/j.1365-2672.2001.01290.x
|
| [96] |
Youssef NH, Duncan KE, McInerney MJ. Importance of 3-hydroxy fatty acid composition of lipopeptides for biosurfactant activity. Appl Environ Microbiol, 2005, 71(12): 7690-7695. DOI:10.1128/AEM.71.12.7690-7695.2005
|
| [97] |
Dehghan-Noude G, Housaindokht M, Bazzaz BS. Isolation, characterization, and investigation of surface and hemolytic activities of a lipopeptide biosurfactant produced by Bacillus subtilis ATCC 6633. J Microbiol, 2005, 43(3): 272-276.
|
| [98] |
Zhao ZZ, Wang QS, Wang KM, et al. Study of the antifungal activity of Bacillus vallismortis ZZ185 in vitro and identification of its antifungal components. Bioresour Technol, 2010, 101(1): 292-297. DOI:10.1016/j.biortech.2009.07.071
|
| [99] |
Shih IL, Kuo CY, Hsieh FC, et al. Use of surface response methodology to optimize culture conditions for iturin A production by Bacillus subtilis in solid-state fermentation. J Chin Inst Chem Eng, 2008, 39(6): 635-643. DOI:10.1016/j.jcice.2008.05.005
|
| [100] |
Davis DA, Lynch HC, Varley J. The application of foaming for the recovery of surfactin from B. subtilis ATCC 21332 cultures. Enzyme Microb Technol, 2001, 28(4/5): 346-354.
|
| [101] |
Glazyrina J, Junne S, Thiesen P, et al. In situ removal and purification of biosurfactants by automated surface enrichment. Appl Microbiol Biotechnol, 2008, 81(1): 23-31. DOI:10.1007/s00253-008-1620-1
|
| [102] |
Gong GH, Zheng ZM, Chen H, et al. Enhanced production of surfactin by Bacillus subtilis E8 mutant obtained by ion beam implantation. Food Technol Biotechnol, 2009, 47: 27-31.
|
| [103] |
Sen R, Swaminathan T. Characterization of concentration and purification parameters and operating conditions for the small-scale recovery of surfactin. Process Biochem, 2005, 40(9): 2953-2958. DOI:10.1016/j.procbio.2005.01.014
|
| [104] |
Sivapathasekaran C, Mukherjee S, Sen R, et al. Single step concomitant concentration, purification and characterization of two families of lipopeptides of marine origin. Bioprocess Biosyst Eng, 2011, 34(3): 339-346. DOI:10.1007/s00449-010-0476-9
|
| [105] |
Wang J, Liu J, Chen H, et al. Characterization of Fusarium graminearum inhibitory lipopeptide from Bacillus subtilis IB. Appl Microbiol Biotechnol, 2007, 76(4): 889-894. DOI:10.1007/s00253-007-1054-1
|
| [106] |
Zhang T, Shi ZQ, Hu LB, et al. Antifungal compounds from Bacillus subtilis B-FS06 inhibiting the growth of Aspergillus flavus. World J Microbiol Biotechnol, 2007, 24(6): 783-788.
|
| [107] |
Velmurugan N, Choi MS, Han SS, et al. Evaluation of antagonistic activities of Bacillus subtilis and Bacillus licheniformis against wood-staining fungi: in vitro and in vivo experiments. J Microbiol, 2009, 47(4): 385-392. DOI:10.1007/s12275-009-0018-9
|
| [108] |
Chen HL, Lee YS, Wei YH, et al. Purification of surfactin in pretreated fermentation broths by adsorptive removal of impurities. Biochem Eng J, 2008, 40(3): 452-459. DOI:10.1016/j.bej.2008.01.020
|
| [109] |
Sabaté DC, Carrillo L, Audisio MC. Inhibition of Paenibacillus larvae and Ascosphaera apis by Bacillus subtilis isolated from honeybee gut and honey samples. Res Microbiol, 2009, 160(3): 193-199. DOI:10.1016/j.resmic.2009.03.002
|
| [110] |
Béchet M, Caradec T, Hussein W, et al. Structure, biosynthesis, and properties of kurstakins, nonribosomal lipopeptides from Bacillus spp. Appl Microbiol Biotechnol, 2012, 95(3): 593-600. DOI:10.1007/s00253-012-4181-2
|
| [111] |
Sang-Cheol L, Kim SH, Park IH, et al. Isolation, purification, and characterization of novel fengycin S from Bacillus amyloliquefaciens LSC04 degrading- crude oil. Biotechnol Bioprocess Eng, 2010, 15(2): 246-253. DOI:10.1007/s12257-009-0037-8
|
| [112] |
Yakimov MM, Timmis KN, Wray V, et al. Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS50. Appl Environ Microbiol, 1995, 61(5): 1706-1713. DOI:10.1128/aem.61.5.1706-1713.1995
|
| [113] |
Kameda Y, Ouhira S, Matsui K, et al. Antitumor activity of Bacillus natto V. isolation and characterization of surfactin in the culture medium of Bacillus natto KMD 2311. Chem Pharm Bull, 1974, 22(4): 938-944. DOI:10.1248/cpb.22.938
|
| [114] |
Hwang YH, Park BK, Lim JH, et al. Lipopolysaccharide-binding and neutralizing activities of surfactin C in experimental models of septic shock. Eur J Pharmacol, 2007, 556(1/2/3): 166-171.
|